Морфометрическая структура прыткой ящерицы обитающей в различных биотопах гомельского района
МИНИСТЕРСТВО
ОБРАЗОВАНИЯ
РЕСПУБЛИКИ
БЕЛАРУСЬ
Учреждение образования
"Гомельский
государственный университет
имени
Франциска Скорины"
Биологический
факультет
Кафедра зоологии и охраны природы
Курсовая работа
Морфометрическая
структура прыткой ящерицы обитающей в различных биотопах гомельского района
Исполнитель:
студент группы Б-52
Казаков Вячеслав Валерьевич
Научный руководитель:
ассистент Кураченко Ирина Витальевна
Рецензент:
доктор медицинских наук, профессор
Калугин Алексей Савельевич
Гомель 2012
Содержание
Введение
Обзор литературы
.1 Ареал прыткой ящерицы
.2 Биологическая характеристика прыткой ящерицы
.3 Плотность населения прыткой ящерицы
.4 Фенотипическое разделение прыткой ящерицы
Объект, программа и методика исследований
Результаты исследований и их обсуждение
.1 Морфофенетические характеристики популяций прыткой
ящерицы, обитающих в Гомельском районе
.2 Особенности структуры популяций прыткой ящерицы в
зависимости биотопа и уровня антропогенной нагрузки
.3 Изучение частоты встречаемости вариантов сочетания щитков
у прыткой ящерицы
Заключение
Список использованных источников
Введение
Оценка современного состояния изучаемой проблемы. До последнего времени
пресмыкающиеся оставались одной из слабоизученных групп, однако в последнее время
началось их активное изучение. Основные задачи исследований по рептилиям
включают следующие положения.
Характеристика современного состояния представителей фауны пресмыкающихся
в Беларуси. Изучается характеристика и экология видов, их морфометрическая и
фенетическая изменчивость, численность, размножение, другие особенности
биологии; оценивается роль пресмыкающихся в биогеоценозах, а также роль
отдельных доминирующих видов в популяциях пресмыкающихся;
Анализ особенностей структуры популяций доминирующих видов, оценка
географической изменчивости ассоциаций рептилий и популяций доминирующих видов.
Выявление основных тенденций динамики герпетокомплексов и популяций
доминирующих видов под влиянием разнотипной антропогенной нагрузки на
окружающую среду;
Разработка рекомендаций по охране и рациональному использованию рептилий
Беларуси.
Актуальность темы: Рептилии как неотъемлемая часть естественных и
трансформированных биоценозов - ценные индикаторы состояния окружающей среды.
Рептилии как одни из наиболее массовых групп животных могут использоваться в
качестве объекта экологического мониторинга, как индикаторы загрязнения
окружающей среды.
Цель работы: изучение
морфо-фенетических признаков (на примере прыткой ящерицы - Lacerta agilis L.) пресмыкающихся юго-востока Беларуси.
Изучение рептилий даёт возможность осуществлять мониторинг состояния их
популяций, предсказывать изменчивость их структуры в связи с изменениями
экологической обстановки и антропогенной нагрузки на биоценозы.
1. Обзор
литературы
1.1 Ареал
прыткой ящерицы
Ареал вида весьма широкий. Она распространена широко в Евразии: от Южной
Англии и центральных районов Франции на восток до Южного Забайкалья,
серо-западного Китая и северной Монголии. Северная граница ареала проходит в
Великобритании, Скандинавии, Швеции до 61° с. ш., не достигая Норвегии. В
Финляндии прыткая ящерица отсутствует. Далее граница проходит через
Волгоградскую и Кировскую области, в Западно-Сибирской низменности поднимается
до 60° с. ш., далее несколько спускается к югу и в Прибайкалье достигает 54° с.
ш. Южная граница ареала проходит по Тувинской АССР севернее хребта Ташу-Ола,
Северо-западному Китаю, заходит в Исыккульскую котловину, через отроги
Джунгарского Алатау, обходит озеро Балхаш с востока. В Казахстане граница идёт
от северного побережья Балхаша, до устья реки Эльбы
[1.20.23]. На Северном Кавказе встречается почти повсеместно [13, 18, 30].
Распространена на Черноморском побережье Кавказа. В Закавказье является горным
видом. По северному побережью Чёрного моря проникает в Крым, но не пересекает
его главную гряду гор [14]. Далее граница доходит до Балканского
полуострова, северо-восточной Турции и северо-западного Ирана [9, 12].
Указанные границы ореала обитания прыткой ящерицы очень приблизительны [4, 6,
7].
В связи с большой протяженностью ареала и различными условиями жизни, вид
распадается на ряд подвидовых форм. Ареал прыткой ящерицы находится в умеренном
и частично субтропическом поясах. Он занимает несколько климатических областей
умеренного пояса (климатологическое районирование по Алисову) [12] Ареал
расположен в климатических поясах, характеризующихся температурой января 0-30°
С, с годовым количеством осадков 100- 2000 мм. Зависимость границы от
температурных условий и солнечной радиации в Европейской части СССР не выявлена.
Среднегодовое количество осадков на большей части ареала не оказывает
существенного влияния на распространение вида. Некоторая зависимость границы
ареала от осадков наблюдается на юге; в Крыму прыткая ящерица не встречается в
местах с относительной влажностью ниже 55% [3]. Восточные и южные популяции
прыткой ящерицы лучше приспособлены к высокогорным условиям, чем западные [8,
19].
В Беларуси прыткая ящерица - самый массовый вид рептилий и водится
повсеместно [21, 23], среди позвоночных изучены мало [25, 26].
Прыткая ящерица обитает на открытых, хорошо прогреваемых солнцем участках
- опушки, поляны и другие типы экотонов в сосновых и других группах лесов.
Предпочитаются в большей степени экотоны сосновых лесов (освоенность 66,2%)
[17].
Поскольку сосновые леса в республике составляют более 50% лесонасаждений,
прыткая ящерицы весьма многочисленна и широко распространена. Она осваивает
также и другие группы лесов, отсутствуя в лесных массивах. Кроме сосняков, в
большей мере предпочитаются экотоны дубрав (освоенность 66,6%), березняков
(30,8%). В еловых лесах отмечена только на участках посадок. Реже всего
встречаются в ольшаниках (7,7%). Луговые биотопы также относятся к обычным
местам обитания прыткой ящерицы (главным образом суходольные, пойменные и речные)
[32].
Нередко прыткая ящерица встречается в типичных для себя биотопах: во
влажных низменных лугах, граничащих с лесами и верховыми болотами; на влажных
участках пойменных озер и рек; на юге республики в жаркие дни она была
обнаружена во влажных ольшаниках. В естественных биотопах, тем не менее,
поселения прыткой ящерицы распространяются весьма мозаично - главным образом по
лесным экотонам, отсутствуя на больших лесных или открытых массивах [7].
Исследования в различных районах Беларуси показали, что в наибольшей
степени ящерицами предпочитается серия антропогенных биотопов, имеющих четко
выраженные экотонные свойства. Это различного рода вырубки (в первую очередь в
сосняках), лесосечные вырубки, просеки квартальные и противопожарные, широкие
трансектальные просеки для ЛЭП, газо- и нефтепроводов и особенно обочины
различного рода дорог (с наибольшим предпочтением откосов и кюветов) [26,29].
Освоенность обочин дорог составляет максимальную величину среди всех
естественных и антропогенных биотопов - 79,4%. Наибольшая предпочитаемость
отдается обочинам дорог (склонам), это объясняется их хорошей прогреваемостью,
незатопляемостью нор. Кроме того, отходы деятельности человека (пищевые
остатки, зерно и другие сельскохозяйственные продукты) привлекают мышевидных
грызунов, которые образуют в придорожной полосе множество нор, используемых
ящерицами. Вместе с тем, различного рода отходы, испражнения
сельскохозяйственных животных, привлекают множество насекомых - основной корм
ящериц.
Можно полагать, что на придорожных полосах создаются наиболее
благоприятные условия для зимовки ящериц: периодически очищаемый с дорог снег,
сгребаемый на обочины, исключает глубокое промерзание почвы [31].
Весьма благоприятны для обитания вида - зоны проведения осушительной
мелиорации. Ящерицы охотно заселяют дамбы польдерных систем и откосные берега
мелиоративных каналов. Особенно четко это прослеживается в пойменных зонах
бассейна реки Припять (регион Полесской низменности) [23].
1.2
Биологическая характеристика прыткой ящерицы
У прыткой ящерицы суженная несколько кпереди голова, длинное,
веретенообразное и гибкое туловище со сравнительно небольшими передними и
задними конечностями и длинным, сужающимся хвостом. Хвост обычно в 1.5 раза
длиннее тела [15].
Голова имеет округленную, пирамидальную форму. На вершине в центре
межчелюстного щитка находится теменной глаз. Глаза расположены по бокам в
средней части головы, развито подвижное третье веко. На конце морды находятся
парные ноздри, а по бокам в задней части головы - вертикальные, овальной формы
слуховые отверстия, затянутые барабанной перепонкой. Туловище удлиненное и
гибкое, отделено от головы шейным перехватом. Граница между туловищем и хвостом
- клоака, которая открывается наружу поперечным отверстием на нижней стороне
тела
Хвост в сечении округлый, постепенно сужающийся к концу. Основание хвоста
у самцов массивнее, чем у самок, из-за расположения там популятивных органов.
Хвост может отсутствовать в результате аутотомии. При регенераци хвоста может
возникнуть двойной хвост.
Конечности сравнительно небольшие, причём задние развиты заметно сильнее
передних. Кисть оканчивается 5 тонкими пальцами, вооруженными коготками [5].
Кожный покров лишен желёз и покрыт роговыми образованиями. Это охраняет
тело от высыхания и механических повреждений. Чешуя покрывает тело. На
неподвижных частях тела находятся роговые чешуйки (щитки), а в местах большой
подвижности - чешуйки-зёрнышки. Часть верхней поверхности головы покрыта
щитками.
Шея от туловища отделяется рядом увеличенных чешуи - воротником. Между
этими рядами и передней частью головы находится горловая складка [7]. Чешуи
спины и боков мелкие, узкие с хорошо выраженными продольными рёбрышками.
Анальный щиток окружен 1-3 рядами преанальных щитков. На внутренней поверхности
бёдер есть ряд чешуек, имеющих в середине отверстие, из которого выделяется
столбик ороговевших клеток - бедренные поры. Они достигают коленного сгиба
(24-26). На хвосте чешуя прямоугольной формы, постепенно уменьшающаяся к концу
хвоста.
В случае аутотомии хвост восстановленный может отличаться по цвету и по
более примитивной чешуе [27, 28].
В пределах Беларуси прыткая ящерица имеет следующие особенности фолидоза
и морфологические характеристики:
· длина тела: 0 до 87,2 мм и 0 до 90 мм;
· длина хвоста: 0 до 121 мм и 0 до 112 мм;
· число чешуи вокруг середины туловища: 0 35 - 45, 0 32-45
· число поперечных рядов брюшных щитков: 0 до 29, 0 до 31.
· число бедренных пор: 0 12-16, 0 11-17;
· число горловой чешуи: 0 16-23, 0 15-24.
В связи с разнообразием мест обитания окраска изменчива, но типичной для
них рисунок из полос и пятен, маскирующий тело при всех условиях, обычно
отличается постоянством [10]. Рисунок на спине у молодых и взрослых ящериц
несколько отличается. Молодь окрашена сверху в коричневые или буровато-серые тона
и на этом фоне у них вдоль хребта проходят 1-2 более тёмные полоски,
окаймленные узкими светлыми линиями. В процессе роста рисунок изменяется.
Тёмные спинные полосы как бы распадаются на отдельные пятна. Нередко они
разделены светлой узкой полосой или же окантованы. Вариант рисунка с тёмными
пятнами и продольными полосами принято считать типичным [16. 18]. Общий фон
окраски верхней части тела у самцов желтовато - бурый, салатный, у самок
белорусских популяций зелёный оттенок отсутствует, преобладает желтовато-коричневый,
коричневато-бурый или буровато-серый цвета. Весной, а иногда и осенью, окраска
особей становится более яркой.
Окраска нижней части тела скромнее; у самок матово-белое брюшко,
желтоватого или голубоватого цвета, у самцов - зеленоватый оттенок. У самцов на
брюхе обычно просматриваются тёмные пятна и крапины. Иногда спинные полосы и
пятна не выражены и рисунок отсутствует. Обоняние развито очень хорошо. Зрение
также хорошо развито и типично для дневного животного. Движения глаз бывают независимы.
Язык используется для осязания. Теменной глаз выполняет определенную
фоторецепторную функцию, повидимому он участвует в поведенческой терморегуляции
[7]. Характерны для прыткой ящерицы колебания температуры тела, от которой
зависит её активность. Максимальная активность наблюдается при 30° С. При
температуре ниже 7.5° С прыткая ящерица впадает в оцепенение [20], может
плавать и проплывать до 10 м, но с обломленным хвостом менее подвижна и не
способна плавать [2].
Избегает повсеместно солодей, солонцеватых и солончаковых почв. На
крайнем северо-западе ареала, в Великобритании обитает на береговых дюнах и
песчаных почвах. В умеренной зоне Европы не избегает песчаных почв с травяной
растительностью. В Восточном Предкавказье уже избегает песчаные участки с
прекционным покрытием травяной растительностью менее 50% [29] и не встречается
в песках Калмыкии, междуречье Волги и Урала. В азиатской части ареала граница
лимитизируется песками зоны пустынь и полупустынь [20, 25].
Убежища прыткой ящерицы делятся на две группы: временные и постоянные
[18, 21]. К временным относятся случайные укрытия (норы земляных ос, мышевидных
грызунов; пространства в кучах камней, отверстия в старых пнях, кучи хвороста и
другие). Постоянные убежища используются систематически в ночной период,
пасмурные дни, во время зимней спячки используются различные по сложности
строения, норы: простые с одним выходом, которые обычно приспособлены для
зимовки, с двумя выходами и лабиринтового типа с несколькими выходами,
используемые в летний период.
Нередко взбирается на кустарники и деревья в поисках пищи [3,13]. Прыткая
ящерица эврифаг. Основу питания составляют насекомые (около 96%), другие мелкие
беспозвоночные (пауки, черви, моллюски, ракообразные, многоножки). Пищевые
объекты невелики. Среди поедаемых объектов встречаются мало подвижные
(гусеницы, моллюски), подвижные (бабочки, жужелицы ), неприятно пахнущие
(клопы), ядовитые и жалящие [18]. Поедает 60% вредных насекомых, что ставит её
в группу полезных животных [29]. После приёма пищи прыткая ящерица ищет хорошо
прогреваемое солнечное место, где греется. Это необходимо для нормальных
процессов жизнедеятельности. В пасмурную погоду движения ящерицы вялые,
предпочитает отсиживаться в норе. Как очень массовый вид служит кормом для
многих видов позвоночных животных. Она была обнаружена в желудках зверей (ёж,
белка, хорь, лисица, норка, куница, барсук, выдра), птиц (белый аист, чайка,
серая цапля, кобчик, полевой лунь, луговой лунь, чёрный и красный коршун,
сарыч, змееяд, ворона, грач, удод, певчий и чёрный дрозд, скворец, сойка),
рептилий (уж, медянка, гадюка, крупные особи своего вида), амфибий (озёрная
лягушка) [11].
К защитным приспособлениям ящерицы относится способность к аутотомии
хвоста. В природных условиях, как среди самцов, так и среди самок наблюдается
иерархия [14].
1.3
Плотность населения прыткой ящерицы
Наибольшая плотность населения прыткой ящерицы отмечается на опушках,
вырубках, свалках, просеках дорог, среди сосняков (до 1000 экз/га). Как видно
из таблицы 1, достаточно высокая плотность может достигаться в экотонах
березняков (до 585 экз/га), лугов (до 300 экз/га), по берегам мелиоративных
каналов (до 700 экз/га). Средневзвешенная плотность (S di ) наиболее высокая для обочин дорог (124,7 экз/га ),
сосняков (77,6 экз/га ), дамб польдерных мелиоративных каналов (51,6 экз/га)
[13].
В целом же по различным типам биотопов плотность вида подвержена
значительной изменчивости [40].
Суточная активность прыткой ящерицы явно зависит от погодных условий,
хотя одним из важнейших приспособлений к их изменчивости служит использование
животными микроклимата [35], что подтверждается и наблюдениями в Беларуси
(например - миграции в жаркое время во влажные биотопы).
Таблица 1 - Плотность населения прыткой ящерицы (Lacerta agilis) и освоенность ею основных типов биотопов [26]
|
Тип биотопа
|
Средневзвешен ная плотность
населения экз/га
|
Количество обследованных
биотопов (n)
|
Освоенность (число
биотопов, в которых отмечен вид, % от исследованных)
|
|
Сосняки
|
77,6 (0 -1000)
|
77
|
66,2
|
|
Ельники
|
19,0 90 - 117,6)
|
19
|
36,8
|
|
Березняки
|
48,2 90 - 585,9)
|
26
|
30,8
|
|
Ольсы
|
0,4 (0 - 9,6)
|
52
|
7,7
|
|
Дубравы
|
46,7 (0 - 140)
|
9
|
66,6
|
|
Луга суходольные и
низинные)
|
31,2 (0 - 300)
|
46
|
47,8
|
|
Луга пойменные (речные)
|
44,8 (0 - 303,6)
|
8
|
25,0
|
|
Луга пойменные (озерные)
|
6,7 (0 - 33,3)
|
5
|
20,0
|
|
Обочины дорог
|
124,7 (0 -600)
|
34
|
79,4
|
|
Мелиоративные каналы
(берега)
|
51,6 (0 - 703)
|
31
|
29,0
|
|
Болота
|
0
|
10
|
0
|
|
Дамбы польдерных
мелиоративных систем
|
69,6 (0 - 100 )
|
5
|
80,0
|
|
Пустоши
|
19,6 (0 - 36,0)
|
7
|
42,8
|
Летом ящерицы начинают появляться из убежищ к 9 часам при температуре
почвы 17 С воздуха в тени - 22 С и на солнце - 28 С. В хорошую погоду к 10
часам, практически все особи поселения покидают норки. Однако в пасмурные дни
ящерицы находятся обычно в убежищах. Например, 27 июля 19979 года в Березинском
заповеднике в 12 часов при температуре почвы и воздуха 15С под камнями
обнаружились малоподвижные самки и самцы. Активность животных сохраняется при
высоких температурах [15, 19].
Конец суточной активности также связан с характером погоды. Повышенная
облачность, дождь во второй половине дня - намного сокращают активный период. В
ясные солнечные дни (май - июнь) ящерицы обычно бывают активны до 19 - 20 часов
(при температуре воздуха в тени 18 С. Но иногда в хорошую погоду после
дождливого периода, мы встречали активных особей после 22 часов. В холодную
солнечную погоду удается отловить до 50% увиденных ящериц, а после дождя
отлавливаются практически все, так как в этом случае подвижность и
маневренность животных явно снижается [39].
Убежища прыткой ящерицы делятся на две группы:
· временные;
· постоянные.
К временным относятся случайные укрытия (норы, землянки ос, мышевидных
грызунов, кучи валежника, бумага и др.). Постоянные убежища используются
систематически (ночной период, пасмурные дождливые дни, зимняя спячка).
Используются различные по сложности строения норы:
· простые с одним выходом;
· имеющие два выхода;
· лабиринтного типа с несколькими выходами.
Сложные норы наиболее эффективны при спасении от хищников. Нередко
убегающая ящерицы спасаются, залезая на деревья (по наблюдениям Пикулика и
других, 1988 на высоту до трёх метров, август, Лоевский район Гомельской
области, опушка дубравы), или же бросаются в воду, где хорошо плавают [11].
Прыткая ящерица - эврифаг. Основу питания составляют насекомые (около 96%),
а также другие мелкие беспозвоночные (пауки, черви, моллюски, ракообразные,
многоножки). Среди поедаемых объектов встречаются как малоподвижные (моллюски,
гусеницы), так и подвижные (жужелицы, бабочки, пчелы, осы, неприятно пахнущие
клоны), ядовитые и жалящие (пчелы и осы). Поедает 60% вредных насекомых, что
ставит ее в группу полезных животных.
После приёма пищи прыткая ящерица ищет хорошо прогреваемое солнцем место,
где греется. Это необходимо для нормальных процессов жизнедеятельности. В
пасмурную погоду движения ящерицы вялые, они предпочитают отсиживаться в норе.
Как очень массовый вид прыткая ящерица служит кормом для многих видов
позвоночных животных. Она была обнаружена в желудках зверей: (ежа, белки,
хорька, лисы, норки, куницы, барсука и выдры), птиц (белого аиста, серой цапли,
полевого луня, чёрного и красного коршуна, змееяда, вороны, грача, удода,
певчего и чёрного дрозда, скворца, сойки), рептилий (ужа, лягушки, гадюки,
крупных особей своего вида), амфибий (озёрной лягушки) [35]. прыткая ящерица популяция биотоп
К защитным приспособлениям относится способность к аутотомии хвоста. В
природных условиях, как среди мужских, так и среди женских особей наблюдается
иерархия [28].
В сентябре - октябре прыткая ящерица уходит на зимовку в зимние норы, как
правило, выходы из нор закрываются листвой и землей, где впадает в зимнюю
спячку. Выход после зимовки в апреле, а на юго-западе Беларуси иногда в конце
марта. Весной у пробудившихся самок имеются мелкие и крупные фолликулы. Их в
яичнике содержится от 21 до 37, в яйцеводах перед кладкой отмечается 4 - 7 яиц.
Самцы по размерам мельче самок. Спариваются в апреле - мае, через месяц после
спаривания в специальной ячейке-ямке, либо в норе перед входом, либо в другом
месте: в рыхлом грунте, обычно на песке, на солнечных местах откладываются
яйца, 2 - 12 штук. Яйца перед кладкой весят 425 - 450 мг. Величина кладки
зависит от размера самки. Инкубация яиц 2 месяца. Появление молоди начинается
не ранее середины июля и может наблюдаться в разных местах в течение августа, в
зависимости от условия развития яиц. Молодое потомство готово при выходе из яиц
к самостоятельному существованию. Половой зрелости достигают к трём годам. [40,
41].
Половая структура популяции в целом по всему ареалу оказывается различной
в разные сезоны. Истинное соотношение полов в популяции, видимо, во все периоды
близко 1/1. Но в некоторых районах ареала наблюдается преобладание численности
женских особей над мужскими среди взрослых животных [26].
Суточные перемещения ящериц невелики, т.к. индивидуальный участок от 15
до 250 см. Может мигрировать из-за засухи, необходимости размножения, половодья
и т.д.
1.4
Фенотипическое разделение прыткой ящерицы
Фен - элементарная вариация признаков или свойств, которая на доступном,
достаточно большом материале не подразделяется [27]. Фены перспективны, по ним
можно выяснить частоту признаков по ареалу [7].
Окраска прыткой ящерицы является фенотипическим признаком. Она
определяется двумя главными компонентами: цветом и рисунком. Сочетание
различных вариантов цвета и рисунка характеризует окраску как весьма
комплексный морфологический признак. При дифференцированном исследовании
составляющих элементов выделяются определенные типы окраски. По фенотипам работ
проведено мало. Экспериментально доказано, что у прыткой ящерицы есть 27
элементарных признаков, которые обуславливают главные элементы рисунка и по ним
можно определить подвид [5]. Подвиды прыткой ящерицы характеризуются также
особенностями фолидоза (щиткование).
Беларусь входит в зону контакта между типичным восточным (Lacerta agilis exigua (Eichwald, 1831 )) и западным (Lacerta agilis agilis (Andrzejowcki, 1832)) подвидами прыткой ящерицы, занимающей обширную
территорию в пределах севере- восточной Украины, Беларуси и восточной Литвы на
западе от Белгородской, Курской, Калужской, юга Московской, Тверской на
востоке. Соотношение признаков восточной и западной форм на протяжении всей
зоны не остаётся постоянным. По мере удаления от основного ареала восточного
подвида признаки этой формы постепенно стушевываются и вытесняются признаками
западной формы.
Этот процесс протекает в направлении с юго-востока на севере- запад. Он
не является равномерным, так как расселение той или иной формы усложняется
наличием различного рода экологических преград и других факторов [19, 20].
Западная географическая группа
1. Прыткая ящерица западная - Lacerta agilis agilis (Andrzejowski, 1758).
2. Прыткая ящерица южная -
Lacerta agilis chersonensis (Andrzejowski, 1832).
3. Прыткая ящерица добруджинская - Lacerta agilis euxinica ( Fuhn et Vancea, 64).
4. Прыткая ящерица боснийская - Lacerta agilis bosnica (Scheiler, 1912).
Подвиды в апреле переходят друг в друга постепенно, и в длинной цепи
замещающих популяций каждая из них представляет собой промежуточную.
Западная географическая группа имеет следующие характеристики (Lacerta agilis (Linnaeus, 1758):
· длина туловища 50,0 - 98,0 мм;
· длина хвоста 0 102- 130 мм и 0 80,0 -138,0 мм;
· индекс хвоста 0 1,24-1,86 и 0 1,18-1,81;
· число чешуи вокруг туловища 0 35-46 и 0 33 - 45.
· число поперечных рядов брюшных щитков 0 25 - 30, О 28 - 32;
· количество бедренных пор 0 11-15, 0 9-15;
· количество горловых чешуй 14-22.
Характер щиткования задненосовой области обычно 1/2 (у большинства
особей), ширина теменной полосы соответствует размерам двух внутренних половин
теменных щитков или 8-14 килеватых чешуй поперёк. Преанальные щитки расположены
в один ряд, в два ряда встречаются редко, только у особей из восточной части
ареала подвида. Анальный щиток у самцов всегда шире, чем у самок [19, 20].
Пилеус и спинка у самцов бурые (до 100%), бока зелёные, у самки - бурые и
в основе рисунка лежат две белые боковые линии, иногда 3 (2 боковые и 1
хребтовая, но сплошная хребтовая полоса отсутствует не менее, чем у 75%).
У самок сплошные полосы на боках отсутствуют у 100% [4, 25]. Южная
географическая группа имеет следующие характеристики (Lacerta agilis chersonensis (Andrzejowcki, 1832)):
· длина туловища 0 68,0-91,0 мм и 0 70 - 94,0 мм;
· длина хвоста 0 98,0-156 мм и 0 84-117,0 мм;
· индекс хвоста 0 1,60-1,75 и 0 1,36-1,54;
· число чешуи вокруг туловища: 36-47;
· число поперечных рядов брюшных щитков 0 26-29 и 0 28-34;
· количество бедренных пор 0 12-19 и 0 13-17;
· количество горловых чешуи 14-25;
· задненосовых щитков 1 -2, чаще 1;
· слуховых 0-2, чаще 1.
Наиболее частая комбинация щитков задненосовой области 1/1 (59%), 1/2
(10,4%) и 2/4 (10%).
Преанальные щитки расположены в 2 ряда, редко в 1 [18, 19].
Тёмная непрерывная полоса спинная по бокам окаймлена светлыми узкими
(боковыми) линиями, которые у 55-90% особей прерывистые. Центральная линия, как
правило, отсутствует (у 75% особей). Достаточно сложные постепенные изменения
признаков и свойств на больших пространствах.
Восточная географическая группа:
1. Прыткая ящерица грузинская - Lacerta agilis grusinica (Peters, 1960).
2. Прыткая ящерица дагестанская - Lacerta agilis boemica (Suchow, 1928).
. Прыткая ящерица короткохвостая - Lacerta agilis brevicandata (Meters, 1958).
. Прыткая ящерица йорская
- Lacerta agilis iorensis ( Peterset Muckhelisknili, 68).
5. Прыткая ящерица восточная - Lacerta agilis exigua (Eichwdld, 1831).
Восточная географическая группа имеет следующие характеристики:
· длина туловища 30,0-114,0 мм
· длина хвоста 36,0-190,5 мм;
· индекс хвоста 0 1,58-2,03 и 0 1,38- 1,68;
· число чешуи вокруг туловища 38 - 54;
· число поперечных рядов брюшных щитков 0 23-32. 0 25-34.
Преанальные щитки обычно расположены двумя полукругами, пара средних
щитков внутреннего круга, как правило, увеличена. Анальный щиток у самцов почти
всегда относительно шире, чем у самок: отношение его к длине 2/2.65.
Задненосовых щитков обычно 2, скуловых тоже 2 (комбинация щитков в
зад-неносовой части обычно 2/2). Широкая тёмная спинная полоса, разделенная
светлой хребтовой линией, занята 13-17 чешуями, которые не выделяются
ребристостью [5, 19]. Пилеус и спина самцов (до 75%) зелёного цвета, самки -
бурого цвета и в основе рисунка лежит 3 непрерывные светлые линии (2 боковых и
1 хребтовая).
Фенотипическим методом исследования можно выяснить меж- и внутрипопуляционную
структуру прыткой ящерицы [28], что косвенно ведёт к выявлению генетической
структуры этих популяций [23]. Фены всегда отражают генетическую структуру
данной особи, а своей частотой генетическую структуру популяции и других групп
особей данного вида [32].
Морфологические признаки (фенотипы и фолидоз) являются одними из исходных
и важных для понимания места в биогеоценозах и для понимания микроэволюционного
процесса в элементарных эволюционных единицах - популяциях данного вида и в их
группах [18, 22].
Такой подход позволяет разрешить противоречие между настоящей
необходимостью генетического исследования огромного числа видов, не затронутых
таким изменением, и невозможностью провести генетическое изучение этих форм
[32, 33].
Это дает ключ к анализу происходящих микроэволюционных событий, скрытых
от кратковременных наблюдений классическими методами /Рис.1/.
2.
Объект, программа и методика исследований
Исследования проводились в летний период 2004 года и летний период 2006
года. Объектом исследований являлись прыткие ящерицы, зарегистрированные в
период практики.
Всего отловлено 118 особей прыткой ящерицы (учтено 220 особей). Собранный
материал фиксировался и хранился в 3% - ном растворе формалина. Отлов ящериц
производили по методике, изложенной в монографии М.М. Пикулика /5/. Камеральная
обработка проводилась в период с августа 2004г. по январь 2007г.
Программа исследований включала:
1. Выбор и описание типичных для
обитания прыткой ящерицы биотопов в зонах с различной антропогенной нагрузкой в
Гомельской области;
2. Отлов прыткой ящерицы в выбранных
биотопах (по три однотипных биотопа в каждой из зон) методом транссекты. Снятие
промеров (длины туловища и длины хвоста) при помощи штангенциркуля. Длина тела
замерялась от кончика носа до анального отверстия, длина хвоста замерялась от
анального отверстия до кончика хвоста;
3. Установление фенетической структуры
популяций прыткой ящерицы;
4. Установление половой структуры
исследуемых популяций;
5. Статистическая обработка и
сравнительный анализ полученных данных.
Систематическое положение:
Тип: хордовые Chordata
Подтип: позвоночные Vertebrata
Раздел: челюстноротые Gnathostomata
Группа: Первичноназемные Amniota
Надкласс: четвероногие Tetrapoda
Класс: пресмыкающиеся (рептилии) Reptilia
Отряд: чешуйчатые Squamata
Подотряд: ящерицы Sauria
Семейство: настоящие ящерицы Lacertidae Bonaparte
Род: ящерицы Lacerta L.
Вид: прыткая ящерица Lacerta agilis L.
Изучение фенов производились по методу А.С. Баранова /1978/ [5],
рассматривались 19 фенов. К ним относятся:
L - сплошные боковые полосы;
L1 - извилистые боковые полосы;
L2 - прерывистые боковые полосы;
D - сплошные хребтовые полосы;
Dd - прерывистая хребтовая полоса;
d - отсутствие хребтовой полосы;
M - наличие спинных пятен;
ММ
- слившиеся спинные пятна;
Мх - спинные пятна пересекают всю
спинную полосу;
Mу - спинные пятна заходят за середину спинной полосы;
M1 - спинные пятна угловатые;
М2 - спинные пятна округлые;
Mm - хаотическое расположение спинных
пятен;
B1 - наличие пятен на каждом срединном брюшном щитке;
В2 -
наличие пятен не на всех срединных брюшных щитках;
В3 - отсутствие пятен на всех срединных брюшных щитках;
В4 -
наличие пятен на каждом боковом брюшном щитке;
B5 - наличие пятен не на всех боковых брюшных щитках;
В6 - отсутствие пятен на всех боковых брюшных щитках.
Все полученные данные обработаны статистически по обычной методике [17,
25].
Характеристика исследованных биотопов:
Микрорайон Волотова
·
Биотоп: обочина
автодороги.
Название ассоциации: дубрава кисличная
Почва: супесчаная
Формула состава древостоя: 8Д1Т1Р
Ярусность древостоя:
А1: Querkus robur L., Acer
platanoides L., Sorbus aucuparia L.
А2: Acer platanoides L.,
Sorbus aucuparia L., Querkus robur L.
А3: Robinia pseudo-acacia L.
Характеристика растений, составляющих покров: Oxalis acetosella L.
Dianthus deltoides L., Dactylis glomerata L., Taraxakum officinale Web.,
Oenothera biennis L., Capsella bursa-pastoris L., Plantago major L.
·
Биотоп: пойменный
луг.
Название ассоциации: суходольный луг
Почва: суглинистая
Характеристика растений, составляющих покров: Centaurea jacea L., Lysimachia nummularia L, Veronika chamaedrys L, Convolvulus arvensis L., Dianthus deltoides L., Polygonum bistorta L., Coronaria flos-cuculi (L.) A. Br., Vicia cracca L., Melilotus albus L., Centaurium vulgare L., Berteroa incana (L.) DC., Knautia arvensis (L.) Coult.
·
Биотоп: сосновый
лес.
Название ассоциации: сосняк черничный
Почва: дерново-подзолистая
Формула состава древостоя: 10С
Характеристика пород, составляющих древесный ярус:
А1: Pinys sylvestris L.
А2: Pinys sylvestris L.
Характеристика растений, составляющих покров: Vaccinium
myrtillus L., Calamagrostis neglecta (Ehrh.) Gaertn., Pteridium aquilinum (L.)
Kuhn, Melampyrum pratense L., Luzula pilosa (L.) Willd., Trientalis europaea L.
Химзавод
·
Биотоп обочина
автодороги
Название ассоциации: сосняк долгомошный
Почва: супесчаная
Формула состава древостоя: 9С1Б
Ярусность древостоя:
А1: Pinys sylvestris L., Betula pendula L.
А2: Pinys sylvestris L., Betula pendula L.3: Rubus idaeus L.
Характеристика растений, составляющих покров: Eriophorum
vaginatum L., Andromeda polifolia L., Carex cinerea Poll., Carex nigra (L.)
Reicherd, Molinia coerulea (L.) Moench.
·
Биотоп: пойменный
луг.
Название ассоциации: суходольный луг
Почва: суглинистая
Характеристика растений, составляющих покров: Vicia cracca L., Melilotus albus L., Melandrium album (Mill.)
Garcke,
Dactylis glomerata L., Ajuga reptans L., Hypericum perforatum L., Centaurium
vulgare L., Berteroa incana (L.) DC., Knautia arvensis (L.) Coult., Dianthus
deltoides L., Coronaria flos-cuculi (L.) A. Br.
·
Биотоп: сосновый
лес.
Название ассоциации: сосняк мшистый
Почва: супесчаная
Формула состава древостоя: 6С4Б
Ярусность древостоя:
А1: Pinus
sylvestris L., Betula pendula Roth, Quercus robur L.
А2: Pinus
sylvestris L., Betula pendula Roth.
А3: Betula pendula Roth,
Quercus robur L.,Frangula alnus Mill.
Характеристика растений, составляющих покров: Melampyrum
pratense L., Vaccinium vitis-idaea L., Pyrola rotundifolia L., Calamagrostis
neglecta (Ehrh.) Gaertn.,
Calamagrostis epigeios (L.) Roth, Chamaecytisus ruthenicus (Fisch. ex
Woloszcz.) Klásková, Veronica officinalis L.
Рекреационная зона “ЧЕНКИ”
·
Биотоп: обочина
автодороги.
Название ассоциации: сосняк мшистый
Почва: песчаная
Формула состава древостоя: 9С1Б
А1
: Pinys
sylvestris L., Betula pendula L.
А2: Pinys sylvestris L.,
Betula pendula L.3: Rubus idaeus L.
Характеристика растений, составляющих покров: Dianthus deltoides L.,
Dactylis glomerata L., Oenothera biennis L., Capsella bursa-pastoris L., Fragaria vesca L., Veronica spicata L., Pteridium aquilinum
(L.) Kuhn., Athyrium filix-femina (L.) Roth., Calluna vulgaris (L.) Hill.,
Carex ericetorum Poll., Agrostis tenuis Sibth.
·
Биотоп: пойменный
луг.
Название ассоциации: суходольный луг
Почва: суглинистая
Характеристика растений, составляющих покров: Centaurea jacea L., Lysimachia nummularia L., Veronika chamaedrys L., Polygonum bistorta
L., Vicia cracca L., Melandrium album (Mill.) Garcke., Dactylis glomerata L.,
Hypericum perforatum L., Centaurium vulgare L., Coronaria flos-cuculi (L.) A.
Br., Convolvulus arvensis L., Dianthus deltoides L.
·
Биотоп: сосновый
лес.
Название ассоциации: сосняк черничный
Почва: песчаная
Формула состава древостоя: 10С
Общее проективное покрытие: 35%
Характеристика растений, составляющих покров: Vaccinium
myrtillus L., Vaccinium vitis-idaea L., Calamagrostis neglecta (Ehrh.) Gaertn.,
Pteridium aquilinum (L.) Kuhn., Melampyrum pratense L., Luzula pilosa (L.)
Willd., Trientalis europaea L., Calluna vulgaris (L.) Hill., Ledum palustre L.,
Polygonatum odoratum (Mill.) Druce. Статистическая обработка данных проводилась по методике,
изложенной Лакиным Г.Ф. /29/.
Основные формулы для расчетов:
Х ср=Sxi /n (2.1)
где: X - среднее значение
xi - варианты
n -
количество вариант
¶2 = S (xi - x)2/ n - 1 (2.2)
Где: ¶2 -
дисперсия
xi - варианта
x -
среднее
n -
количество вариант
mx = √ 1/ n-1 (Sxi2/n - x) (2.3)
Где: mx - ошибка средней
n -
количество вариант
xi - варианта
x - среднее
tф = x1 - x2 / √ m1 - m2 (2.4)
При: p = 0.95; tт = 1.96; tф
< tт
Где: tф - нормированное отклонение
x1
- средняя для
первой выборки
x2
- средняя для второй
выборки
m1
- ошибка средней
для первой выборки
m2
- ошибка средней
для второй выборки
p -
точность исследований
tт
- критерий точности
3.
Результаты исследований и их обсуждение
3.1
Морфофенетические характеристики популяций прыткой ящерицы, обитающих в
Гомельском районе
Исследования морфофенетической структуры популяций прытких ящериц,
проведенные нами в трех типовых биотопах Гомельского района, дают основание
считать, что различия этих популяций не очень значительны. Анализ данных по
методу Т-стандартного свидетельствует о том, что дифференцировка средних
морфофенетических параметров рассматриваемых популяций неслучайна, и
обусловлена различиями структуры однотипных биотопов. По результатам наших
исследований средние величины длины тела и длины хвоста ящериц (таблица 2), отловленных в биотопе обочина
автодороги, составили 74.5±11.1 и 102.3±15.5 мм
соответственно. При этом, минимальная длина тела составляла 53 мм (1
экземпляр), максимальная - 91 мм (3 экземпляра); минимальная длина хвоста - 69
мм (1 экземпляр), максимальная - 123 мм (1 экземпляр). Выборка из биотопа 20
особей. Фен D обнаружен у 15 особей (75 %), фен Dd - у 10 особей (50 %), фен Dc - у 17 особей (85 %), фен Dm - у 15 особей (75 %), фен L - у 16 особей (80 %), фен Ld - у 14 особей (70 %), фен M - у 18 особей (90 %), фен Mm - у 13 особей (65 %), фен MM - у 14 особей (70 %), фен Mi - у 11 особей (55 %), фен Md - у 17 особей (85 %), фен B3 - у 13 особей (65 %).
Средние величины длины тела и длины хвоста ящериц, отловленных в биотопе
сосновый лес, составили 78.1±11.2 и 112.8±16.2 мм
соответственно. При этом минимальная длина тела составляла 57 мм (1
экземпляр), максимальная - 90 мм (1 экземпляр); минимальная длина хвоста - 75
мм (1 экземпляр), максимальная - 132 мм (1 экземпляр). Выборка из биотопа - 20
особей. Фен D обнаружен у 15 особей (75 %), фен Dd - у 10 особей (50 %), фен Ds - у 11 особей (55 %), фен Dn - у 13 особей (65 %), фен L - у 17 особей (85 %), фен Ld - у 18 особей (90%), фен M - у 16 особей (80%), фен Mm - у 10 особей (50 %), фен MM - у 6 особей (30 %), фен Mo - у 16 особей (60 %), фен Md - у 13 особей (65 %), фен B3 - у 17 особей (85 %).
Средние величины длины тела и длины хвоста ящериц, отловленных в биотопе
пойменный луг, составили 71.4±11.1 и 116.8±16.6 мм
соответственно. При этом минимальная длина тела составляла 55 мм (2
экземпляра), максимальная - 90 мм (1 экземпляр); минимальная длина хвоста - 82
мм (2 экземпляра), максимальная - 141 мм (1 экземпляр). Выборка из биотопа - 20
особей. Фен D обнаружен у 15 особей (75 %), фен Dd - у 12 особей (61 %), фен Ds - у 1 особи (фен Dc - не обнаружен), фен Dn - у 1 особи (фен Dm - не обнаружен), фен L - у 19 особей (95 %), фен Ld - у 17 особей (85 %), фен M - у 14 особей (75 %), фен Mm - у 14 особей (75 %), фен MM - у 6 особей (30 %), фен Mo - у 13 особей (65 %), фен Mx - у 14 особей (75 %), фен B2 - у 17 особей (85 %), фен B4; B5 - у
18 особи (90 %).
Таким образом, морфометрические и фонетические параметры прытких ящериц,
обитающих в различных биотопах Гомельского района, различны. Для анализа
различий морфометрических параметров ящериц из различных биотопов мы применили
методику Т-стандартного (Лакин Г. Ф. /29/), при расчетах использовалась формула
2.4. В качестве нулевой гипотезы принято, что различия морфометрических
параметров ящериц из различных биотопов носят случайный характер. Однако
величины tф, полученные нами, удовлетворяют условию tф >tт, следовательно, нулевая гипотеза неверна (tф 2,07 - 4,21). Таким образом, различие
морфометрических и фонетических параметры прытких ящериц, обитающих в различных
биотопах Гомельского района неслучайны. Различия фенов популяций, обитающих в
различных биотопах, проявляются незначительно для ящериц, отловленных на
обочине автодороги и в сосновом лесу (темная и светлая срединные полосы
соответственно), отличие ящериц, отловленных на пойменном лугу, состоит в
отсутствии срединной линии и множественных пятнышках на брюшных щитках (у
остальных пятнышки отсутствуют). По остальным индикаторным фенам все три
выборки одинаковы, различия - лишь в процентном соотношении особей с
противоположными индикаторными фенами.
Таблица 2 - Средние морфофонетические параметры прытких ящериц, обитающих
в биотопах Гомельского района
|
Параметр
|
X±m
|
|
Биотоп обочина
автодороги
|
|
Длина тела (мм)
|
74.5±11.1
|
|
Длина хвоста (мм)
|
102.3±15.5
|
|
Средний индикаторный фен
|
D;Dd;Dc;Dm;L;Ld;M;Mm;Mi;Md;B3
|
|
Биотоп сосновый лес
|
|
Длина тела (мм)
|
78.1±11.2
|
|
Длина хвоста (мм)
|
112.8±16.2
|
|
Средний индикаторный фен
|
D;Ds;Dn;Dd;L;Ld;M;Mm;Mo;Md;B3
|
|
Биотоп пойменный луг
|
71.4±11.1
|
|
Длина хвоста (мм)
|
116.8±16.6
|
|
Средний индикаторный фен
|
L;Ld;d;M;Mm;Mo;Mx;B2;B4;B5
|
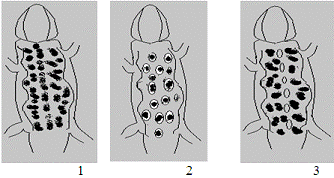
1- обочина автодороги
2 - пойменный луг
3 - сосновый лес
Рисунок 1 - Средние индикаторные фены прытких ящериц, занимающих
различные биотопы Гомельского р-на
Морфофонетические параметры прытких ящериц, обитавших в биотопах
Гомельского района, полученные нами в 2004 г., приведены в таблице 3. Для
анализа данных мы применяли методику двухфакторного дисперсионного анализа
(Рокицкий П.Ф. /30/), схема которого описана в приложении. В качестве факторов
влияния на фонетическую структуру популяции прыткой ящерицы нами
рассматривались погодные условия июня и июля месяцев 2004 года, а также общие
условия биотопов. Данные метеорологических условий взяты нами по сводкам
объединения “Гомельоблгидромет”.
Таблица 3 - Средние данные морфофонетических параметров прытких ящериц,
занимающих различные биотопы Гомельского р-на
|
июнь 2004 год
|
|
Биотоп
|
Обочина Автодороги
|
Сосновый лес
|
Пойменный луг
|
|
Длина тела (мм)
|
73.6±10.7
|
81.6±13.2
|
73.8±10.7
|
|
Длина хвоста (мм)
|
88.5±13.1
|
99.6±13.3
|
84.8±12.2
|
|
Индикаторный фен
|
D;Ds;Dn;Dd;L;Le;M;Mm;Mi;Mx;B2;B4
|
D;Ll;M;Mm;MoMx;B2;B4
|
D;Dc;Dm;L;Ll;M;Mm;B3
|
|
июль 2004 год
|
|
Биотоп
|
Обочина Автодороги
|
Сосновый лес
|
Пойменный луг
|
|
Длина тела (мм)
|
74.5±11.1
|
78.1±11.2
|
71.4±11.1
|
|
Длина хвоста (мм)
|
102.3±15.5
|
112.8±16.2
|
116.8±16.6
|
|
Индикаторный фен
|
L;Ld;d;M;Mm;Mi;MoB2;B4
|
D;Ds;Dn;Dd;L;Ld;MMm;Mo;Md;B3
|
L;Ld;d;M;Mm;
Mo;Mx;B2;B4;B5
|
|
tф
|
Длина тела
|
1,30
|
1,84
|
1,10
|
|
Длина хвоста
|
2,53
|
2,19
|
2,78
|
Нами установлено, что наибольшее влияние оказывает совместное действие
этих факторов (85.96%), в меньшей степени сказывается воздействие условий
биотопов (13.76%) и еще меньше влияние метеоусловий (0.33%), т.е. воздействие
метеоусловий на фонетику популяций опосредовано биотопом обитания.
Статистическая обработка и последний анализ материалов по фонетике
прыткой ящерицы обследованных биотопов показали, что не все выделенные фены
достаточно показательны.
В результате отбора для анализа были оставлены три фена, по которым можно
проследить популяционные различия:
·
фен D, характеризующий сплошную хребтовую
полосу;
·
фен L, показывающий прерывистость боковых
полос;
·
фен Mm, который обозначает хаотическое
расположение спинных пятен.
А - Окрестности химзавода им. М. В. Ломоносова
Биотоп: А1 сосновый лес.
Нами получены следующие частоты фенов по биотопу А: для самок (учтено 28
особей) - фен D - 100%, Dd - 0%, Dc -
46%, Dn - 54%, L - 100%, Ll -
54%, M - 100%, Mm - 54%, MM -
0%, Mn - 46%, Mo - 38%, Md-
69%, B3 - 81%; для самцов (отловлено 20
особей) - фен D - 100%, Dd - 0%, Dc - 58%,
Dn - 42%, L - 100%, Ll -
46%, M - 100%, Mm - 29%, MM -
0%, Mn - 71%, Mo - 25%, Md-
25%, B3 - 25%. Таким образом, частоты фенов
для самцов и самок прыткой ящерицы, обитающих в биотопе сосновый лес в
окрестностях промышленного предприятия, существуют значительные различия в
частотах встречаемости некоторых фенов (Mm, Mn, Md, B3), и, следовательно, различны средние индикаторные фены: ♂
- D, Dc, Dm, L, Ld, M, Mn, Mi, Mx, B2, B4; ♀ - D, Ds, Dn, L, Ll, M, Mm, Mi, Md, B3.
Биотоп: А2 пойменный луг.
Частоты фенов следующие: для самок (учтено 24 особи) - фен D - 67%, Dd - 71%, Dc -
56%, Dn - 81%, L - 71%, Ll -
41%, M - 75%, Mm - 28%, MM -
28%, Mn - 44%, Mo - 44%, Md -
39%, B3 - 71%; для самцов (отловлено 16
особей) - фен D - 89%, Dd - 65%, Dc -
61%, Dn - 61%, L - 73%, Ll -
74%, M - 85%, Mm - 45%, MM -
32%, Mn - 23%, Mo - 41%, Md-
68%, B3 - 35%. Значительные отличия в
частотах наблюдаются для фенов Ll, Mn, B3 и Md.
Соответственно средние индикаторные фены: ♂ - D, Dd, Dc, Dn, L, Ll, M, Mm, Mo, Md, B2, B4; ♀ - D, Dd, Dc, Dn, L, Ld, M, Mn, Mi, Mx, B3.
Биотоп: А3 обочина автодороги.
Частоты фенов следующие: для самок (учтено 24 особи) - фен D - 100%, Dd - 58%, Dc -
33%, Dn - 4%, L - 71%, Ll -
41%, M - 79%, Mm - 0%, MM -
100%, Mn - 0%, Mo - 42%, Md -
74%, B3 - 71%; для самцов (отловлено 20
особей) - фен D - 100%, Dd - 62%, Dc
-38%, Dn -46%, L - 85%, Ll -
18%, M - 81%, Mm - 0%, MM -
100%, Mn - 0%, Mo - 19%, Md -
86%, B3 - 11%. Значительные отличия в
частотах наблюдаются для фенов Dn, Ll, B3 и Mo.
Соответственно средние индикаторные фены: ♂ - D, Dd, Ds, Dm, L, Ld, M, MM, Mi, Md, B2, B4; ♀ - D, Dd, Ds, Dm, L, Ld, M, MM, Mi, Md, B3.
В - дачная зона “Кленки”.
Биотоп: B1 сосновый лес.
Нами получены следующие частоты фенов по биотопу А: для самок (учтено 20
особей) - фен D - 100%, Dd -57%, Dc -
82%, Dn - 100%, L - 89%, Ll -
32%, M - 100%, Mm - 0%, MM -
43%, Mn - 57%, Mo - 18%, Md-
82%, B3 - 57%; для самцов (отловлено 20
особей) - фен D - 100%, Dd - 23%, Dc -
77%, Dn - 100%, L - 96%, Ll -
36%, M - 100%, Mm - 0%, MM -
45%, Mn - 55%, Mo - 23%, Md-
82%, B3 - 27%. Таким образом, сильно
различаются частоты фенов: Dd и B3 , следовательно, различны средние
индикаторные фены: ♂ - D, Dc, Dn, L, Ld, M, Mn, Mi, Md, B2, B4; ♀ - D, Dd, Dc, Dn, L, Ld, M, Mn, Mi, Md, B3.
Биотоп: В2 пойменный луг.
Частоты фенов следующие: для самок (учтена 31 особь) - фен D - 19%, Dc - 26%, Dn -
26%, L - 100%, Ll - 35%, M -
100%, Mm - 100%, Mo - 61%, Md -
26%, B3 - 55%; для самцов (отловлено 15
особей) - фен D - 21%, Dc - 21%, Dn -
21%, L - 100%, Ll - 32%, M -
100%, Mm - 100%, Mo - 37%, Md-
42%, B3 - 32%. Значительные отличия в
частотах наблюдаются для фенов Mo и B3. Соответственно средние
индикаторные фены: ♂ - d, L, Ld, M, Mm, Mi, Mx, B2, B4; ♀ - d, L, Ld, M, Mm, Mo, Mx, B3.
Биотоп: B3 обочина автодороги.
Частоты фенов следующие: для самок (учтено 23 особи) - фен D - 100%, Dd - 56%, Dc -
62%, Dn - 75%, L - 88%, Ll -
41%, M - 100%, Mm - 16%, MM -
84%, Mn - 0%, Mo - 69%, Md -
19%, B3 - 59%; для самцов (отловлено 11
особей) - фен D - 100%, Dd - 61%, Dc
-55%, Dn -61%, L - 95%, Ll -
44%, M - 100%, Mm - 33%, MM -
67%, Mn - 0%, Mo - 33%, Md -
89%, B3 - 0%. Значительные отличия в
частотах наблюдаются для фенов Mo, Md, B3. Соответственно средние индикаторные фены: ♂ - D, Dd, Dc, Dn, L, Ld, M, MM, Mi, Md, B2, B4; ♀
- D, Dd, Dc, Dn, L, Ld, M, MM, Mo, Mx, B3.
С - рекреационная зона в урочище “Ченки”.
Биотоп: B1 сосновый лес.
Нами получены следующие частоты фенов по биотопу: для самок (учтено 16
особей) - фен D - 96%, Dd -100%, Dc -
96%, Dn - 96%, L - 39%, Ll -
23%, M - 100%, Mm - 93%, MM -
0%, Mn - 7%, Mo - 29%, Md-
33%, B3 - 68%; для самцов (отловлено 22
особи) - фен D - 86%, Dd - 100%, Dc -
87%, Dn - 77%, L - 59%, Ll -
18%, M - 100%, Mm - 95%, MM -
0%, Mn - 5%, Mo - 64%, Md-
59%, B3 - 23%. Таким образом, сильно различаются
частоты фенов: L, Mo, Md, B3 ,
следовательно, различны средние индикаторные фены: ♂ - D, Dd, Dc, Dn, L, Ld, M, Mm, Mo, Md, B2, B4; ♀
- D, Dd, Dc, Dn, l, M, Mm, Mi, Mx, B3.
Биотоп: C2 пойменный луг.
Частоты фенов следующие: для самок (учтено 22 особи) - фен D - 56%, Dd - 100%, Dc -
100%, Dn - 100%, L - 53%, Ll -
65%, M - 100%, Mm 100%, Mo -
78%, Md - 16%, B3 - 84%; для самцов (отловлено 18 особей) - фен D - 72%, Dd - 92, Dc -
100%, Dn - 92%, L - 55%, Ll -
55%, M - 89%, Mm - 100%, Mo -
59%, Md- 29%, B3 - 44%. Значительные отличия в частотах наблюдаются для фена
B3, для других - разница невелика.
Соответственно средние индикаторные фены: ♂ - D, Dd, Dc, Dn L, Ll, M, Mm, Mo, Mx, B2, B4; ♀
- D, Dd, Dc, Dn L, Ll, M, Mm, Mo, Mx, B3.
Биотоп: C3 обочина автодороги.
Частоты фенов следующие: для самок (учтено 20 особей) - фен D 96%, Dd - 0%, Dc -
77%, Dn - 81%, L - 43%, Ll -
25%, M - 100%, Mm - 93%, MM -
0%, Mn - 7%, Mo - 29%, Md -
32%, B3 - 68%; для самцов (отловлено 12
особей) - фен D - 86%, Dd - 0%, Dc
-89%, Dn -84%, L - 41%, Ll -
18%, M - 100%, Mm - 68%, MM -
0%, Mn - 32%, Mo - 54%, Md -
59%, B3 - 14%. Значительные отличия в
частотах наблюдаются для фенов Mo, Md, B3. Соответственно средние индикаторные фены: ♂ - D, Dc, Dn, l, M, Mm, Mo, Md, B2, B4; ♀ - D, Dc, Dn, l, M, Mm, Mi, Mx, B3.
В результате исследований биотопов окрестностей УНБ “Чёнки” в 2006 году,
получены следующие результаты. Рассмотрим некоторые характеристики фенетической
структуры исследуемых популяций. Биотоп сосновый лес. Выборка из биотопа
- 9 особей. Средний индикаторный фен: D / Dc / L / M / MM / B4. Фен D
обнаружен у 5 особей (55,6 %), Dd - 3
особей (33,4 %), Dc - 6 особей
(66,7 %), Ds - 2 особей (22,3 %), d - 1 особи (11,2 %), L - 6 особей (66,7 %), Le - 2 особей (22,3 %), Ll - 1 особи (11,2 %), M - 9 особей (100 %), Mo - 2 особей (22,3 %), Mx - 1 особи (11,2 %), MM - 5 особей (55,6 %), Mm - 1 особи (11,2 %), B4 - 6 особей (66,7 %).
Биотоп обочина автодороги. Выборка из биотопа - 11 особей. Средний индикаторный
фен: D / Ds / L / M / Ml / B3. Фен D обнаружен у 5 особей (45,4 %), Dd - 3 особей (27,2 %), Dc - 2 особей (18,1 %), Ds - 6 особей (54,5 %), d - 3 особей (27,2 %), L - 6 особей (54,5 %), Le - 1 особи (9 %), Ll - 2 особей (18,1 %), l - 2
особей (18,1 %), M - 11 особей (100
%), Mo - 5 особей (45,4 %), Ml - 6 особей (54,5 %), B3 - 6 особей (54,5 %).
3.2
Особенности структуры популяций прыткой ящерицы в зависимости биотопа и уровня
антропогенной нагрузки
Произвольно нами было выделено три зоны с различным уровнем антропогенной
нагрузки: А - окрестности промышленного предприятия
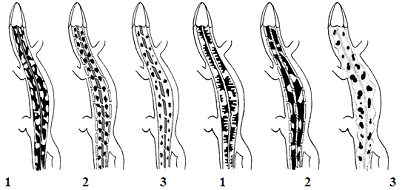
1 - окрестности УНБ “Чёнки”; 2 -
окрестности д. Клёнки; 3 - окрестности д. Цыкуны
А - биотоп сосновый лес; Б - биотоп обочина
дороги
Рисунок 2 - Средние индикаторные фены популяций прыткой
ящерицы, обитающих в различных биотопах Гомельского района (УНБ «Ченки») в
летний период 2006 г.
Средние индикаторные фены прытких ящериц, обитающих в различных биотопах,
различны для зон с разной антропогенной нагрузкой, типов биотопов и пола
особей. Для подтверждения этого факта мы подвергли полученные нами данные
трехфакторному дисперсионному анализу (Лакин, /30/). В качестве факторов
влияния на фенетическую структуру популяций мы рассматривали антропогенную
нагрузку (фактор А), тип биотопа (фактор В) и пол особей прыткой ящерицы
(фактор С), а также совместное действие этих факторов. Полученные нами
результаты анализа свидетельствуют о том, что сила влияния различных факторов
на различные фены неодинакова. Данные анализа представлены в таблице 4.
Таблица 4 - Сила влияния различных факторов на частоту фенов в популяциях
прыткой ящерицы
|
Сила влияния факторов, %
|
|
Фен
|
A
|
B
|
C
|
AB
|
AC
|
BC
|
ABC
|
Z
|
|
D
|
7.23
|
27.35
|
--
|
37.78
|
--
|
--
|
--
|
27.64
|
|
Dd
|
--
|
5.02
|
--
|
68.11
|
5.16
|
5.19
|
--
|
16.52
|
|
Dc
|
24.39
|
--
|
--
|
--
|
5.76
|
--
|
--
|
69.85
|
|
Dn
|
47.21
|
15.93
|
--
|
4.09
|
--
|
9.62
|
--
|
23.15
|
|
L
|
19.44
|
18.73
|
--
|
12.75
|
--
|
--
|
--
|
49.08
|
|
Ld
|
--
|
--
|
--
|
--
|
--
|
--
|
--
|
100
|
|
M
|
26.53
|
8.11
|
--
|
13.41
|
--
|
--
|
--
|
51.95
|
|
Mm
|
41.51
|
18.14
|
--
|
21.95
|
--
|
--
|
--
|
18.40
|
|
MM
|
25.59
|
34.61
|
4.21
|
27.35
|
2.25
|
--
|
--
|
5.99
|
|
Mn
|
23.74
|
24.99
|
--
|
25.83
|
--
|
--
|
--
|
25.44
|
|
Mo
|
16.68
|
--
|
--
|
13.73
|
5.26
|
--
|
--
|
64.33
|
|
Md
|
8.31
|
11.37
|
4.71
|
16.43
|
--
|
4.63
|
--
|
54.55
|
|
B2, B4
|
--
|
5.14
|
57.70
|
--
|
--
|
--
|
--
|
37.16
|
Примечание:
А - антропогенное воздействие;
В - тип биотопа;
С - пол;
АВ, АС, ВС, АВС - совместное воздействие соответствующих факторов;
Z - не
учитываемые в исследовании факторы.
Из таблицы 4 видно, что максимальное влияние уровень антропогенной
нагрузки оказывает на частоту проявления фенов Dn и Mm, а
ее влияние на фены Dd, Ld и B2,B4 статистически
не доказано. Влияние условий биотопа сильно сказывается на частотах фенов MM, D и Mn и
статистически не доказано для фенов Dc, Ld и Mo. Фен B2, B4 (множественные черные пятна на
брюшных щитках) является половым признаком - частота проявления на 57,7%
зависит от пола особи; очень незначительно влияние пола на фены Md и MM менее 5%, для остальных фенов влияние пола не доказано. Что
касается совместного действия факторов, то наиболее велика роль антропогенной
нагрузки и типа биотопа; так, фен Dd на 68% зависит от воздействия суммы этих факторов - т. е. является
зависимым от антропогенной нагрузки и типа биотопа. Отсутствие статистической
зависимости частоты фенов от совместного влияния трех факторов (АВС)
объясняется, на наш взгляд, множественным характером зависимости между ними.
Тем же, вероятно, объясняется и отсутствие влияния на фен Ld (наличие извилистой боковой полосы).
Сравнивая данные по морфофенетическим показателям прыткой ящерицы
окрестностей УНБ «Ченки» за летний период 2004 и 2006 года, можно утверждать,
что популяции стабильны. По основным индикаторным фенам отличий не выявлено
(исключение составляет наличие пятен на брюшных щитках, половой признак).
3.3
Изучение частоты встречаемости вариантов сочетания щитков у прыткой ящерицы
При изучении популяций прыткой ящерицы обращалось большое внимание на
изменчивость фолидоза, т.е. количество и распределение щитков на определенных
участках тела. Наибольшее систематическое значение имеют щиткование
задненосовой и преанальной областей.
По характеру щиткования задненосовой области наличие в первом ряду одного
щитка характерно для западного типа прыткой ящерицы, а двух щитков - для
восточного типа.
Как видно из таблицы 5 во всех изученных популяциях прыткой ящерицы
имеются варианты и западного и восточного типов, т.е. характер щиткования
смешанный.
При этом соотношение отдельных вариантов в каждом типе в разных
популяциях колеблется в широких пределах, поэтому целесообразно было
рассматривать суммарные значения вариантов по количеству щитков, т.е.
соотношение двух типов щиткования.
Во всех трёх популяциях преобладает западный тип (с 1 щитком в первом
ряду) - около 65%. Здесь наиболее сильно проявляются признаки подвида L. a. Chersonensis (55,5%). Восточный тип щиткования (с
2 щитками в первом ряду) встречался во всех трёх популяциях в примерно
одинаковом соотношении - около 23-25%.
Соотношение количества полных рядов прианальных щитков в изученных
популяциях различно, но во всех трёх преобладали ящерицы с 2 рядами среди
ящериц пойменного луга - 62,6%, обочина автодороги - 61,4% и сосновый лес -
60,8%. Отнесение выборок по этому признаку к какому-либо из рассматриваемых
типов затруднено, так как наличие двух рядов присуще и для восточных подвидов и
для западных. Несмотря на это, можно говорить о том, что в исследуемых популяциях
преобладает западный тип прыткой ящерицы с выраженным подвидом L. a. chersonensis.
Таблица 5 - Частота встречаемости различных вариантов сочетания
задненосовых щитков и числа полных рядов прианальных щитков (в %)
|
Характеризующие параметры
|
Расположение популяций
|
|
|
Пойменный луг
|
Обочина автодороги
|
Сосновый лес
|
|
Щиткование задненосовой
области, N a/Lor
|
1/1
|
55,5±5,0
|
52.2±4.0
|
48,8±3.0
|
|
1/2
|
17,8±3,5
|
18,2±2.5
|
20,2±2,0
|
|
2/1
|
17,8±3.5
|
18.0±2,5
|
17,2±2,0
|
|
2/2
|
8,9±0,3
|
9,2±0,4
|
8,6±0,3
|
|
Количество полных рядов преанальных
щитков, Pr, an
|
1
|
38,6±0,5
|
40,2±0,6
|
36,8±0.5
|
|
2
|
61,4±0,6
|
62,6±0.6
|
60,8±0.5
|
Таким образом, в исследуемых биотопах отмечено преобладание западного
типа L. a. agilis.. В популяциях пойменного луга более сильно
проявляются признаки подвида L. a. chersonensis, у которых комбинация 1/1 достигает
55,5%.
По соотношению количества полных рядов преанальных щитков популяции не
имеют различий. Отнести к какому-либо типу подвидовой формы исследуемые
популяции трудно, так как, наличие 2-х рядов характерно как для южного подвида L. a. chersonensis, так и для восточного - L. a. eligua.
Заключение
В ходе исследований отмечено преобладание западного типа L. a. agilis. В исследуемых биотопах Гомельского района. В
популяциях пойменного луга более сильно проявляются признаки подвида L. a. chersonensis, у которых комбинация 1/1 достигает
55,5%.
По соотношению количества полных рядов преанальных щитков популяции не
имеют различий. Отнести к какому-либо типу подвидовой формы исследуемые
популяции трудно, так как, наличие 2-х рядов характерно как для южного подвида L. a. chersonensis, так и для восточного - L. a. eligua.
Установлены следующие морфофенетические параметры для популяций прыткой
ящерицы: обочина автодороги: длина тела - 74.5±11.1
мм, длина хвоста - 102.3±15.5 мм, средний
индикаторный фен - D;Dd;Dc;Dm;L;Ld;M;Mm;Mi;Md;B3; сосновый лес: длина тела - 78.1±11.2 мм, длина хвоста - 112.8±16.2 мм, средний индикаторный фен - D;Ds;Dn;Dd;L;Ld;M;Mm;Mo;Md;B3; пойменный луг: длина тела - 71.4±11.1 мм, длина хвоста - 116.8±16.6
мм, средний индикаторный фен - L;Ld;d;M;Mm;Mo;Mx;B2;B4;B5.
Максимальное влияние уровень антропогенной нагрузки оказывает на частоту
проявления фенов Dn и Mm, а ее влияние на фены Dd, Ld и B2,B4 статистически не доказано. Влияние
условий биотопа сильно сказывается на частотах фенов MM, D и Mn и статистически не доказано для
фенов Dc, Ld и Mo.
Фен B2, B4 (множественные черные пятна на брюшных щитках) является
половым признаком - частота проявления на 57,7% зависит от пола особи; очень
незначительно влияние пола на фены Md и MM менее 5%, для остальных фенов
влияние пола не доказано. Что касается совместного действия факторов, то
наиболее велика роль антропогенной нагрузки и типа биотопа; так, фен Dd на 68% зависит от воздействия суммы
этих факторов - т. е. является зависимым от антропогенной нагрузки и типа
биотопа.
Список
использованных источников
1 Алисов Б.П. Палтараус Б.В. Климатология. - М.:
Издательство Московского университета, 1974. - 297 с.
2 АлисовБ.П. Климат СССР. -М.: Высшая школа, 1969.- 104
с.
Банников А.Г., Даревский И.Е., Рустамов А.К., и др.
Земноводные и пресмыкающиеся СССР.-М.: Просвещение: 1971. - 303 с.
Банников А.Г., Даревский И.Е., Ищенко В.Г. и др.
Определитель замноводных и пресмыкающихся фауны СССР.- М.: Просвещение. 1977.- 414
с.
Бахарев В.А. К вопросу о подвидовой структуре
прыткой ящерицы в Белоруссии: Вопросы герпетологии.-Л.: Наука, 1981. - 215 с.
Бахарев В.А. Географические особенности
распределения численности пресмыкающихся Белоруссии // Всесоюзная
зоогеографическая конференция. -М.: 1984. - С.273
Бобров В. Ящерицы по имени ящурки. // Наука и жизнь.
- 1998.-№ 5. -С.158.
Энцыклапедыя прыроды Беларусi: Бясхвостыя земнаводныя - Мiнск: 1983. Т.1. - 390 с.
Герасимов В.П. Животный мир нашей родины.-М.:
Просвещение, 1965.- 126 с.
Гаранин В.И. Амфибии и рептилии в питании
позвоночных. Природные ресурсы Волжско-Камского края.- Казань., 1976.-выпуск 4
- 86 с.
Дарафееу А. I iнш. Рыбы, земнаводныя, паузуны. - Мн.: Народная асвета, 1981.
-159 с.
Даревский С.И. Очередные задачи советских
герпетологов. Вопросы герпетологии // Ш Всесоюзная герпетологическая
конференция, 1973.- М.: Просвещение, 1973 -С. 75-76.
Деккерт Г., Деккерт К. Как ведут себя животные. -
М.: Лесная промышленность, 1985. - 57 с.
Дмитриев Ю. Земноводные пресмыкающиеся. - М.: Олимь,
1998. - 303 с.
Карр А. Рептилии. - М.: Наука, 1975. - 190 с.
Курскова Г.Н., Пикулик М.М. О морфофизиологических
адаптациях амфибий Белоруссии. - Мн.: Навука, 1976. - 227 с.
Наумов Н.П., Карташёв Н.Н. Зоология позвоночных. - М.:
Высшая школа, 1979. - ч.2. - 333 с.
Новожёнов Ю.И., Береговой В.Е., Хохуткин И.Н.
Обнаружение границ элементарных популяций полиморфных видов по частоте
встречаемости форм. Проблемы эволюции. - Новосибирск.: Наука, 1973. - С. 252 -
261.
Пикулик М.М., Бахарев В.А., Косов Е.А.
Пресмыкающиеся Белоруссии. - Мн.: Наука и техника, 1988. - 165 с.
Пикулик М.М. Ящарка. - Мн.: Наука и техника, 1997. -
№5 - С. 185.
Прыткая ящерица. Монографическое описание вида /
Институт биологии развития им. Н.К. Кольцова. Под ред. Яблокова А.В. - М.:
Наука,1976. - 374 с.
Пикулик М.М. Они нужны природе.- Мн.: Наука и
техника, 1984. - 127 с.
Сапожников Ю.Ф. Материалы к изучению амфибий и
рептилий Белоруссии. Фауна и экология наземных позвоночных Белоруссии. Мн.:
Навукова думка, 1978. - 94 с.
Терентьев П.В., Тертышников М.Ф. Прыткая ящерица как
консумент и продуцент. - М.: Наука, 1976. - 302 с.
Тимофеев - Ресовский Н.В., Яблоков А.В., Глотов Н.В.
Очерк учения о популяции. - М.: Наука. 1973. - 278 с.
Терентьев П.В.. Чернов С.А. Определитель
пресмыкающихся и земноводных. - М.: Советская литература, 1949. - 340 с.
Турупина А.В., Подмарев В.И. Территориальное
распределение фенов и выделение генетико-пространственных группировок в
популяции прыткой ящерицы. Физиологическая и популяционная экология животных. -
Саратов.:Изд-во Саратовского университета, 1978. - С. 68 -71.
Щепотьев А.В. К вопросу о хозяйственном значении
прыткой ящерицы в полезащитных лесопарках. - Зоология животных, 1952. - Т. 31.
- №4. - С.574 - 582.
Щербак Н.Н. Земноводные и пресмыкающиеся. -
Симферополь.: Крым, 1966. - 59 с.
Щербак Н.Н., Осташко Н.Г., Даревский И.С.
Монографическое описание вида. - М.: Наука. 1976. - 374 с.
Яблоков А.В. Популяционная биология. - М.: Наука,
1987.- 302 с.