Особенности опыления подсолнечника при свободном цветении и в условиях сетчатого изолятора
Дипломная
работа
Особенности
опыления подсолнечника при свободном цветении и в условиях сетчатого изолятора
Введение
Тенденция расширения посевных площадей под
подсолнечником (Helianthus
annuus L.)
связана, в первую очередь, с тем, что в последние годы культура оказалась в
числе наиболее рентабельных в системе растениеводства [Кривошлыков, 2005]. Во
многих районах Западного Предкавказья данная сельскохозяйственная энтомофильная
культура является одной из самых высеваемых. К тому же подсолнечник в
агроландшафтной системе южных регионов России в летний период считается ведущим
медоносным растением, привлекающим насекомых-опылителей. Тесная приуроченность
пчелиных к питанию на посевах цветущих энтомофильных культур позволяет
рассматривать опылителей, как один из важнейших компонентов полезной биоты,
населяющей эти культуры [Ченикалова, 2005].
Селекционная работа с масличным подсолнечником в
СССР ведется около 60 лет. За это время советскими селекционерами (М. Плачек,
В. С. Пустовойтом, Л. А. Ждановым, В. И. Щербина и др.) достигнуты выдающиеся
успехи в создании высокоценных сортов данной культуры.
Исключительное значение в селекции подсолнечника
имеют работы во Всероссийском научно-исследовательском институте масличных
культур (ВНИИМК), в котором их более 60 лет возглавлял академик В. С.
Пустовойт. Сорта института и его сети занимают свыше 90% [Подсолнечник.
Достижения селекции … , 2012].
Целью данной работы является изучение
особенностей опыления подсолнечника при свободном цветении и в условиях сетчатого
изолятора. Для достижения поставленной цели решились следующие задачи:
изучение общих характеристик подсолнечника, как
масличной культуры;
изучение особенностей его опыления и главных его
опылителей;
изучение особенностей селекции подсолнечника в связи
с проблемой пчелоопыления;
изучить особенности опыления подсолнечника на
центральной экспериментальной базе ВНИИМК.
Данная тема в настоящее время является
актуальной, в связи с использованием подсолнечника как основной масличной
культуры.
Работа излагается на 51 странице, состоит из
введения, трех глав, заключения, списка используемых источников, включающего 40
наименований, и одного приложения. Также работа включает 4 таблицы и 12
рисунков.
1. Обзор литературы
.1 Общая характеристика
подсолнечника
Подсолнечник - основная масличная культура в
нашей стране [Подсолнечник, 1975]. В бывшем СССР ежегодно выращивалось свыше
5,3 млн. т семян подсолнечника, вырабатывалось около 2 млн. т подсолнечного
масла, что составляет три четверти общего производства пищевых растительных
масел. В настоящее время выращивается: в РФ - около 6 млн. т (на 4,5 млн. га),
в ЮФО - более 3 млн. т (на 2,5 млн. га), в Краснодарском крае - около 1,1 млн.
тонн (на 550 тыс. га) [Малюга, Квашин, Загорулько, 2011].
Основной, ценной в хозяйственном отношении
частью урожая подсолнечника, ради которой он и возделывается, являются семена
[Подсолнечник, 1975], из которых и производят масло. Подсолнечное масло имеет
высокие пищевые и вкусовые достоинства. Оно используется непосредственно в пищу
и широко применяется в различных отраслях пищевой промышленности. В масле
содержатся биологически активные вещества - фосфатиды, жирорастворимые витамины
и провитамины A, D,
E [Никитчин, 2002].
Содержание токоферолов (витамин Е) в масле достигает 60-80%, фосфатидов
(фосфолипидов) - 0,7-1,0%, из которых 55-65% приходится на долю лецитинов -
веществ, наиболее ценных для пищевых и технических целей. При разработке семян
на масло вырабатывается 33-35% шрота (жмыха), в котором содержится до 40%
протеина. Этот протеин характеризуется довольно высоким содержанием незаменимых
аминокислот [Дьяков, 2004].
Подсолнечный шрот широко используется в
животноводстве в качестве высококонцентрированного белкового корма. Он является
важным белковым компонентом при производстве различных комбикормов.
Подсолнечник широко используют как силосную
культуру, особенно в районах Нечерноземья и Восточной Сибири.
Среди полевых культур трудно найти другую,
которая была бы так щедра, как подсолнечник. Один гектар его посева при
урожайности семян 25ц/га дает 12 ц масла, 8 ц шрота (3 ц белка), 5 ц лузги (0,7
ц дрожжей), 15 ц корзинок (10 ц хорошего сена), 25-30 кг меда и много другой
нужной продукции [Малюга, Квашин, Загорулько, 2011].
Подсолнечник имеет мощный, облиственный,
зеленый, травянистый, в нижней части одревесневший стебель, заканчивающийся
соцветием. Поверхность стебля шероховатая, матовая, опушена многоклеточными
волосками. Узлы стебля открытые, по своему строению мало отличаются от
междоузлий. На поверхности стебля под листьями выступают ниспускающиеся из
черешков жилки, образующие характерную угловатость стебля, которая выражена тем
сильней, чем благоприятнее условия питания и водоснабжения растений. Стебли
растений селекционных сортов масличного подсолнечника не только не ветвятся, но
в пазухах их листьев даже не закладываются в почки [Володарский, Газенко,
1960].
После цветения верхняя часть стебля поникает в
направлении на восток. Длина стебля варьирует от 60 см у скороспелых до 200 см
и более у растений силосных сортов. В верхней части диаметр стеблей
подсолнечника в 2 и более раз меньше, чем у основания.
Растения подсолнечника имеют обычно хорошо
выраженный главный стержневой корень. В отличие от стебля корни подсолнечника
многократно ветвятся [Пустовойт, 1967].
Листья у подсолнечника простые, черешковые, без
прилистников. Расположены на стебле спирально и только самые нижние -
супротивно. Растения сортов среднеспелой группы имеют обычно по 27-30 листьев.
Длина и ширина листьев в зависимости от их яруса и условий внешней среды могут
меняться от нескольких сантиметров до 0,5 м. Пластинки листьев простые,
цельные, цельнокрайние у самых нижних и с зубчатыми или крупнопильчатыми краями
у всех остальных листьев.
Цветки подсолнечника собраны в соцветие -
многоцветковую верхушечную корзинку, имеющую форму круглого плоского, выпуклого
или вогнутого диска. Корзинка окружена оберткой из нескольких рядов листочков.
Максимальное число цветков, закладывающихся в корзинке и после оплодотворения
превращающихся в семянки, может достигать 8 тыс. Но в большинстве случаев
корзинка имеет от 1,2 тыс. до 3-4 тыс. цветков. Диаметр корзинок культурного
подсолнечника зависит от условий произрастания и варьирует от 10 до 26 см.
Для корзинок характерно также положение на
стебле. К моменту зацветания корзинки почти прямостоящие и слегка наклонены
набок, позднее, при наливании семян, они принимают различное положение - от
прямостоящих до опрокинутых.
Дифференциация цветочных бугорков идет от края
корзинки к центру: наружные образуют язычковые цветки, остальные - трубчатые.
Этот этап органогенеза характеризуется для подсолнечника процессами микро- и
мегаспорогенеза (в пыльниках формируется пыльца, в завязи - зародышевый мешок).
На последующих этапах происходит рост язычковых
и трубчатых цветков в длину. Вместе с околоцветником растут тычиночные нити и
пестик, пыльники желтеют, разворачивается обертка корзинки [Подсолнечник ,
1975].
Трубчатые цветки - обоеполые. К составным частям
трубчатого цветка относятся: чашечка, венчик, тычинки и пестик.
Чашечка состоит из двух раздельных чашелистиков,
сильно редуцированных, имеющих вид маленьких рожков, длиной 3-5 мм, светлой или
темнофиолетовой окраски. Прикреплены чашелистики к верхнему концу завязи в
месте соединения ее с венчиком и легко обламываются.
Венчик - правильный, сростнолепестный и имеет
форму трубочки. Нижняя часть трубочки венчика несколько сужена и образует
вздутие в виде кольца, с внутренней стороны которого находится ткань, покрытая
железками, выделяющими нектар. Верхняя же часть ее более расширена и заканчивается
пятью зубчиками яйцевидной формы. Окраска венчика светложелтая, коричневая или
темнофиолетовая. Размер венчика в длину 8-12 мм и в ширину (в диаметре) 2-5 мм.
Число тычинок 5. Нити их свободны, а пыльники,
сросшиеся в одну общую трубочку, окружающую столбик пестика. Трубочка пыльников
цилиндрической формы, несколько суженная к концам и имеет гофрированную
поверхность темносерого или темнофиолетового цвета. Размер трубочки в длину 3-7
мм и в диаметре 1-2 мм. Верхний конец ее заканчивается пятью створочками,
раскрывающимися при выходе пыльцы и рыльца. При созревании пыльников их гнезда
вскрываются в полость трубочки продольными щелями и вся трубочка заполняется
пыльцой.
Количество пыльцы в пыльниках у различных форм и
сортов подсолнечника разное. Пыльцевые зерна подсолнечника в воздушном
состоянии имеют овальную форму, окрашены большей частью в желтый или оранжевый
цвет и покрыты острыми шипиками. Во влажной среде пыльцевые зерна очень быстро
набухают и принимают шаровидную форму.
Пестик состоит из двух плодолистиков. Основными
частями его являются: завязь, столбик и рыльце.
Рыльце разделяется на две ветви или лопасти
длиной в 3-5 мм. Трех и больше лопастные рыльца чаще всего наблюдаются у
цветков центральной части корзинки. Лопасти окрашены в желтый и коричневый
цвет, с наружной стороны покрыты ворсинками, направленными вверх.
Столбик - округлой формы, внутри имеет
канальчик. При полном выходе рыльца из пыльниковой трубочки столбик достигает
8-12 мм длины и около 1 мм толщины.
Завязь - нижняя, одногнездная, содержит одну
обратную семяпочку. Во время раскрытия цветка завязь имеет размер в длину 7-10
мм и ширину 3-5 мм [Подсолнечник, 1940].
Особенность цветков подсолнечника
состоит в том, что они имеют нектарники, которые выделяют нектар. Пчелы,
собирая нектар, опыляют цветки. Выделение нектара зависит от температуры
окружающего воздуха и запаса влаги в почве. Наиболее благоприятной температурой
считается 20- 25 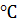 [Гаврилова, Анисимова, 2003].
[Гаврилова, Анисимова, 2003].
Процесс цветения. Бутонизация
подсолнечника происходит в период наиболее интенсивного его роста. Зацветание
подсолнечника характеризуется появлением язычковых цветков корзинки, что
наблюдается обычно в утренние или вечерние часы дня. Одновременно с этим
происходит довольно интенсивный рост цветоложа в ширину. Через 1-2 дня в
зависимости от условий погоды, язычковые цветки полностью раскрываются. Вслед
за раскрытием язычковых цветков, еще имеющих скученные и не вполне
расправившиеся лепестки, по краю корзинки начинают зацветать трубчатые цветки.
Цветение трубчатых цветков
происходит отдельными поясами, чередующимися друг за другом в направлении от
края корзинки к ее центру. У сортов масличного подсолнечника число их доходит
до 8-10 и иногда больше. В свою очередь количество раскрывшихся цветков в
отдельном поясе также значительно колеблется, в зависимости от места
расположения их в корзинке и условий погоды.
К концу цветения корзинки происходит
завядание язычковых цветков, которые довольно долго сохраняют свою свежесть,
пока не отцветут последние трубчатые цветки в корзинке.
Развитие цветка. Образование и
развитие трубчатых цветков в прицветниках начинается еще задолго до начала
цветения корзинки. С раскрытием листовой обертки и язычковых цветков можно видеть,
как цветоложе сплошь покрыто молодыми бутонами различного размера. Более
крупные из них расположены ближе к периферии корзинки, менее крупные в средней
части ее и мелкие - в центре корзинки.
Зацветание трубчатых цветков
начинается с раскрытия бутонов, что начинается рано утром. Еще с вечера
предыдущего дня бутоны крайнего пояса корзинки начинают увеличиваться в
размерах. Затем на верхушках бутонов образуются пять зубчиков, одновременно с
этим, под влиянием роста тычиночных нитей, начинается выход пыльников. В
результате зубчики все больше расходятся в стороны и, наконец, бутон
преобразуется в трубочковидный венчик цветка. Выход пыльников продолжается в
течение 1-1,5 часа. Как только пыльники поднимутся своей нижней частью до
уровня зубчиков венчика, дальнейший рост тычиночных нитей прекращается и вместе
с этим заканчивается выход пыльников. К этому времени пыльники вполне уже
созревают и их трубочка заполнена зрелой пыльцой. В момент полного выхода
пыльников из венчика рыльце с сомкнутыми лопастями находится между тычиночными
нитями под трубочкой пыльников, так как столбик пестика за это время очень мало
увеличивается в росте. Через некоторое время начинается ускоренный рост
столбика пестика, благодаря чему рыльце выходит в пыльниковую трубочку и постепенно
выталкивает оттуда пыльцу. В это время створочки пыльников открываются и на
поверхность выступает пыльца. Выход пыльцы очень заметен по ее ярко-желтой
окраске. Пыльца, выступившая из пыльников, разносится насекомыми и ветром и
через 2-3 часа ее почти не остается на пыльниках.
После выхода пыльцы у раскрывшихся
цветков в течение дня заметных изменений не происходит, наступает как бы период
покоя. И только к 4-5 часам вечера снова начинается рост столбика пестика:
рыльце выходит из пыльниковой трубочки наружу и одновременно выносит оттуда
остатки пыльцы. Через 1-2 часа рыльце полностью выходит из пыльников, вслед за
этим происходит постепенное расхождение его лопастей, после чего рыльце вполне
подготовлено к восприятию пыльцы. При полном выходе рыльца начинается увядание
тычинок, нити их сокращаются и пыльники постепенно сползают по столбику пестика
вовнутрь венчика. Такое разновременное развитие половых органов цветка, при
котором вначале созревают мужские органы, а потом женские, в биологии принято
обозначать специальным термином - протерандрией.
Рыльце в отношении выполнения своих
функций находится в несколько иных условиях, чем тычинки. Оно прежде всего
зависит от опылителей, переносчиков на него пыльцы, затем от возможности
прорастания пыльцы и оплодотворения ею семяпочки завязи данного цветка. Средняя
продолжительность жизненного цикла трубчатого цветка от раскрытия бутона и до
завядания рыльца, при нормальных условия оплодотворения, равна 1,5 суток
[Арнольдова, 1982].
Несколько последовательных фаз в развитии
цветка подсолнечника, с которыми приходится иметь дело во время искусственных
скрещиваний:
1) начало раскрытия бутона между 3-4 часами
утра;
) выход пыльников между 4-6 часами утра;
) выход пыльцы между 6-8 часами утра;
) выход рыльца между 5-7 часами вечера;
) раскрытие лопастей рыльца между 7-9
часами вечера;
) увядание рыльца в первой половине следующего
дня [Подсолнечник, 1940].
1.2 Процесс опыления у подсолнечника
В отношении способа опыления у подсолнечника в
литературе имеются различные указания. Одни авторы указывают, что подсолнечник
является типичным перекрестноопылителем и самостерилен, другие считают, что,
наряду с перекрестным опылением, у подсолнечника имеет место самоопыление и
самофертильность.
Так, например, Фрувирт говорит, что самоопыление,
достигаемое посредством изолирования всей корзинки, хотя и дает семянки, но эти
семянки являются щуплыми [Фрувирт, 1914].
Шулл указывает, что он, ведя в течение четырех
лет генетические работы по подсолнечнику, ни разу не получил семянок, путем
самоопыления.
Вагнер, изучая самоопыление и автостерильность у
различных видов подсолнечника, пришел к заключению, что популяции H.
аnnuus L.,
подобно другим перекрестноопылителям, как рожь и свекла состоит из
самоопыляющихся и автостерильных типов, из числа которых могут быть выделены
соответсвующие автофертильные формы. По его опытам, изолированные корзинки, в
среднем, дали 12,7 урожая [Подсолнечник … , 1940].
Плачек Е.М., на основании долголетнего
инцухтирования подсолнечника, приходит к выводу о наличии биотипов с высокой
автофертильностью и что самоопыление подсолнечника передается по наследству
[Плачек, 1930].
Пустовойт В.С. указывает, что при изоляции целых
корзинок процент оплодотворения у различных биотипов колеблется от 0 до 90%
[Пустовойт, 1928].
Названные авторы установили наличие самоопыления
у подсолнечника путем искусственного изолирования корзинок в период их
цветения. Однако, такое самоопыление нельзя отождествлять с возможным
самоопылением в естественных условиях, т.е. у открыто цветущих растений, так
как самоопыление в условиях искусственной изоляции происходит под известным
«принуждением». Если мы искусственно создаем растениям такую среду, при которой
они лишаются возможности получить чужую пыльцу (с других растений), то они
биологически вынуждены будут частично самооплодотвориться.
Проведенный сотрудниками ВНИИМК опыт [Научный
отчет ВНИИМК … , 1936] показывает, что у подсолнечника имеет место избирательна
способность оплодотворения. В строении трубчатого цветка подсолнечника и
биологии его развития имеется особая приспособленность к перекрестному
опылению. Во-первых, цветение всей корзинки и отдельного ее цветка происходит
совершенно открыто (цветки открытовенчиковые), благодаря чему, рыльца вполне
доступны для чужой пыльцы. Во-вторых, происходит равномерное созревание тычинок
и рыльца в цветке (явление протерандрии), что ведет к исключению возможного
самоопыления в пределах цветка. В-третьих, имеет место физиологическая
невосприимчивость рыльца к своей пыльце.
Проведенные наблюдения над самоопылением в
пределах корзинки, группы цветков и отдельного цветка, путем применения
искусственной изоляции, показывают, что при изоляции целых корзинок или группы
цветков происходит частичное завязывание семян от самоопыления (7-9 %) и
совершенно остаются неоплодотворенными завязи у индивидуально изолированных
цветков. Исходя из этого, можно предполагать, что завязывание семян в
изолированных корзинках происходит в результате переопыления цветков в пределах
корзинки, что свободно может происходить в силу встряхивания корзинки ветром,
осыпания пыльцы с верхних цветков на рыльце нижних, соприкосновения рылец с
пыльниками соседних цветков и т.д., но не от самоопыления у отдельно взятого
цветка.
Таким образом следует, что подсолнечник является
ксеногамным (перекрестноопыляющимся) растением, но, наряду с ксеногамией, у
него может иметь место гейтеногамия. Последнее происходит, главным образом, в
силу отсутствия или недостаточности перекрестного опыления [Подсолнечник,
1940].
1.3 Опылители подсолнечника
Перекрестное опыление у подсолнечника
осуществляется, главным образом, насекомыми и частично ветром. Поэтому,
подсолнечник относится к числу энтомофильных растений.
Среди насекомых, как опылителей подсолнечника, в
наших условиях наибольше значение имеют пчелы. При цветении подсолнечника в
поле ежедневно можно наблюдать массовое посещение цветущих корзинок пчелами,
особенно в утренние часы дня, при выходе пыльцы из пыльников. Пчелы прилетают
сюда за нектаром и пыльцой. Для сбора нектара они последовательно обыскивают
цветки и одновременно с этим собирают на свои лапки и брюшко большое количество
пыльцы, легко прилипающей к опушенным частям тела насекомых. Очень часто можно
видеть, как пчелы от пыльцы буквально становятся желтыми (рисунок 2).
Помимо пчел, подсолнечник посещают в довольно
большом количестве и другие насекомые, которые также способствуют перекрестному
переопылению его цветков. Подсолнечник - это хороший, распространенный медонос
с широкой экологической пластичностью. Вызревание полноценных семян подсолнечника
невозможно без опыления его насекомыми, основными из которых являются
одиночные, медоносные пчелы и шмели.
Для определения эффективности опыления был
проведен опыт с изоляторами, который показал, что корзинки подсолнечника,
находящиеся в контроле, т.е. в условиях свободного опыления, отличались хорошей
выполненностью семянок и их плотным прилеганием друг к другу. Корзинки с
изоляторами имели мелкие, не плотно прилегающие друг к другу семянки.
Перелетая с одного растения на другое, в поисках
нектара, они, вместе с этим, переносят пыльцу на рыльца цветков. Полезность
пчел при опылении подсолнечника и этим самым повышение его урожайности доказана
[Богоявленский, 1935].
Таким образом, самоопыление в пределах одной
корзинки носит ярко выраженные признаки инцеста, поэтому энтомофильное опыление
цветков современных сортов является веским аргументом в пользу организации
планового перекрестного опыления этой культуры [Голиков, 2008].
Значение ветра в опылении подсолнечника
находится в известной зависимости от того, как часто он может быть в период
цветения растений, какой силы или скорости, направления и, наконец,
приспособленности самой пыльцы к переносу ветром.
Ветер, как агент перекрестного опыления у
подсолнечника, имеет относительно большое значение при переопылении растений в
пределах одного и того же массива, чем в разных массивах, удаленных друг от
друга на 200-250 м. Поэтому, при решении вопроса об установлении
экспериментально обоснованной пространственной изоляции, для посевов разных
сортов подсолнечника необходимо считаться, прежде всего, с наличием
насекомых-опылителей и, в первую очередь, пчел, способных перелетать на
довольно значительное расстояние. Отсюда возникает вопрос - способны ли пчелы
одновременно посещать цветущий подсолнечник на разных массивах, расположенных
друг от друга на расстоянии 1-2 км, так как с этим связана пригодность
применяемых в настоящее время норм пространственной изоляции для подсолнечника
[Подсолнечник, 1940].
Созревание полноценных семян подсолнечника
невозможно без опыления его насекомыми, основными из которых являются
одиночные, медоносные пчелы и шмели.
Изучение опылителей подсолнечника показало, что
одним из важных факторов аттрактивности, от которой зависит посещаемость
подсолнечника пчелиными, является интенсивность выделения нектара. Численность
пчелиных на подсолнечнике, помимо нектаропродуктивности последнего, зависит
также от сорта культуры, степени цветения и времени суток.
Также, проведенный опыт показал, что корзинки
подсолнечника, находящиеся в контроле, т.е. в условиях свободного опыления,
отличались хорошей выполненностью семянок и их плотным прилегание друг к другу.
Корзинки с изоляторами имели мелкие, не плотно прилегающие друг к другу семянки
(таблица 1).
Таким образом, самоопыление в пределах одной
корзинки носит ярко выраженные признаки инцеста, поэтому энтомофильное опыление
цветков современных сортов является веским аргументом в пользу организации
планового перекрестного опыления этой культуры [Голиков, 2008].
Пчелиные (Apoidea)
составляют одно из крупнейших надсемейств отряда перепончатокрылые (Hymenoptera)
и в настоящее время насчитывают около 21 тыс. видов, принадлежащих к 520 родам
6 семейств.
Пчелы - это энтомофильные насекомые и посещают
практически все покрытосеменные растения, пыльца и нектар которых служит для
них трофическим субстратом. Как известно, первоначальным этапом антофилии у
пчел являлась поллинофагия (питание пыльцой). Впоследствии нектар в качестве
дополнительного компонента для привлечения насекомых вошел в состав пищи пчел.
Питание пыльцой, а затем нектаром сыграло важную роль в эволюции пчел и
повлияло на их морфологию, биологию и поведение. Корм для личинок пчелиные
обычно заготавливают в виде смеси пыльцы и нектара, соотношение которых
варьирует у разных видов. Отдельные виды пчел добавляют в «хлебец» секреты
мандибулярных желез, обладающие фунгицидным и ингибирующим действием [Пчелы,
2012].
Таблица 1 - Заполненность корзинок семянками у
различных сортов подсолнечника при изоляции и свободном опылении [Голиков,
2008]
|
Вид
изолятора
|
Общее
число семянок в корзинках, шт.
|
Выполненных
семянок, %
|
|
ВНИИМК
80
|
Кавказец
|
ВНИИМК
8931 улучшенный
|
ВНИИМК
80
|
Кавказец
|
ВНИИМК
8931 улучшенный
|
|
Капронове
изоляторы
|
1175±11,0
|
1178±13,2
|
1128±13,1
|
3,3±0,51
|
6,3±0,66
|
2,4±0,45
|
|
Марлевые
изоляторы
|
1123±18
|
1127±1,9
|
1118±13,1
|
0,6±0,23
|
1,3±0,32
|
0,6±0,19
|
|
Контроль
|
1215±1,9
|
1239±4,1
|
1244±3,6
|
96,1±0,59
|
96,1±0,55
|
96,3±0,53
|
Пчелы - это энтомофильные насекомые и посещают
практически все покрытосеменные растения, пыльца и нектар которых служит для
них трофическим субстратом. Как известно, первоначальным этапом антофилии у
пчел являлась поллинофагия (питание пыльцой). Впоследствии нектар в качестве
дополнительного компонента для привлечения насекомых вошел в состав пищи пчел.
Питание пыльцой, а затем нектаром сыграло важную роль в эволюции пчел и
повлияло на их морфологию, биологию и поведение. Корм для личинок пчелиные
обычно заготавливают в виде смеси пыльцы и нектара, соотношение которых
варьирует у разных видов. Отдельные виды пчел добавляют в «хлебец» секреты
мандибулярных желез, обладающие фунгицидным и ингибирующим действием [Пчелы,
2012].
По типу трофических связей пчел разделяют на
полилектов и олиголектов. Самки полилектов собирают пыльцу с широкого круга
растений - это широкие полилекты или только с некоторых видов растений разных
семейств - узкие полилекты. Олиголекты питаются на цветках преимущественно
одного семейства - широкие олиголекты или исключительно одного семейства.
Иногда пыльца собирается только с цветков растений одного или нескольких близких
родов - узкие олиголекты или даже с растений одного вида - монолекты.
В условия Кубани обилие цветущей растительности
способствует широкому распространению полилектичных видов, численность которых
составляет около 60% от всех пчелиных.
Несмотря на пищевую специализацию, при
отсутствии своих кормовых растений олиголекты могут выкармливать потомство
пыльцой других растений. Отродившиеся у таких видов пчел личинки при наличии
специализированных растений собирают пыльцу с них, а не с тех растений, из
которых был сформирован хлебец, что предполагает возможность передачи
трофических привычек пчел с генами [Голиков, 2009].
Медоносная пчела (Apis
mellifera
L.). Медоносную
пчелу как наиболее многочисленного опылителя подсолнечника рассмотрим более
подробно.
Пчелы относятся к надсемейству летающих
насекомых отряда перепончатокрылых, родственному осам и муравьям. Пчелы
трудолюбивые медоносные насекомые, которое состоит из трех подвижно соединенных
отделов - голова, грудь, брюшко.
Голова пчелы - передняя часть, из крепкого и
плотного хитина, в виде округленно треугольной формы. На голове пчелы
сосредоточена большая часть органов чувств, из-за наличия двух боковых сложных
и трех простых теменных глазиков, помимо этого, еще многочисленное количество
усиков, антенн. В нижней передней части головы находятся верхние челюсти. Также
у пчелы есть хоботок, который отличается сложным строением. С его помощью
рабочие особи собирают крошечные капельки нектара с цветущих растений.
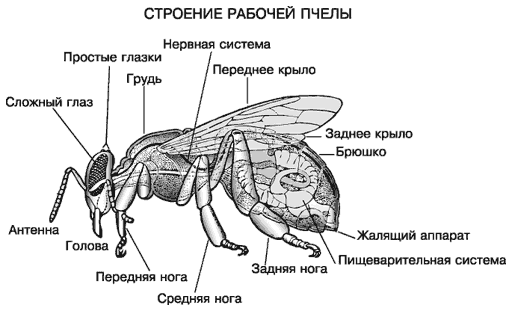
Строение рабочей пчелы [Строение рабочей … ,
2012]
Второй отдел - грудь пчелы - соединен с головой
тонкой хитиновой пленкой, которая не сковывает движение и позволяет насекомому
смотреть во все стороны. Можно разделить грудь на четыре соединенных части,
одна из которых входит в состав брюшка. Передние и задние пары перепончатых
крыльев прикрепляются к отделам груди, также как и членистые ножки. С помощью
прочных жилок и тонкой пленки на крыльях, пчела может превосходно летать и
держаться в воздухе. Если насекомому не нужно передвигаться, то крылья
складываются одно на другое вдоль тельца. В случае же, если пчела планирует
взлететь, крылья расправляются, при помощи крючочков соединяются и образуют с
двух сторон, как бы, одно большое крыло. У пчелы очень мощные и развитые мускулы.
Пчела легко преодолевает большие расстояния, перемещаясь с одного цветущего
поля на другое. На каждой членистой ножке пчелы есть два небольших коготка и,
так называемая, подушечка. Коготки необходимы насекомому для передвижения по
шероховатой поверхности, а с помощью подушечек, пчела присасывается к гладкому
покрытию, чтобы не скользить. У медоносной пчелы на ножках размещаются
маленькие корзиночки. В них насекомое собирает цветочную пыльцу для
последующего переноса в улей.
В брюшке располагаются все самые главные
жизненно важные органы пчелы, а так же кровеносная, пищеварительная и
дыхательная системы. Брюшко пчелы окрашено в желтый цвет и разделено кольцами,
которые увеличиваются или уменьшаются при дыхании, или когда зобик наполняется
нектаром.
Пчелиная матка. Матка в семье является
единственной полноценной самкой с хорошо развитыми половыми органами. От неё
происходит весь состав семьи: рабочие пчёлы, трутни и молодые матки. Матка
постоянно находится в окружении рабочих пчёл, которые ухаживают за ней.
Присутствие в семье матки пчёлы узнают по её запаху. Матка выделяет особое
вещество так называемое «маточное вещество», которое слизывают рабочие пчёлы из
окружающей её «свиты». Матка выделяется среди рабочих пчёл более крупными
размерами тела от 18 до 25 мм, а у рабочих пчёл от 12 до 15 мм, и сравнительно
короткими крыльями (по отношению к длине тела). По сравнению с рабочими пчёлами
хоботок у матки короче (3,5 мм). Она, как и рабочие пчёлы, имеет жало, но
использует его лишь в борьбе с другими матками. Плодная матка летом весит
примерно 0,25 г, вес же рабочей пчелы равен в среднем 0,1 г. Органы размножения
у пчелиной матки развиты очень сильно. Яичники состоят из 180-200 яйцевых
трубочек; в них зарождаются и развиваются яйца. От яичников отходят парные
яйцеводы, соединяющиеся в один непарный яйцевод, с которым небольшим
семяпроводящим каналом соединён семяприёмник.
При осеменении маток сперма трутней, содержащая
огромное количество сперматозоидов, попадает в семяприёмник матки, где и
хранится в течение её жизни. Если при этом в созревшие яйца из семяприёмника
проникнут сперматозоиды (по 8-12 штук), то яйца будут оплодотворены. Если же
сперма трутней на них не попадёт, яйца останутся неоплодотворёнными.
Следовательно, матка откладывает яйца оплодотворённые и неоплодотворённые. Из
неоплодотворённых яиц развиваются только самцы-трутни. Из оплодотворённых яиц
развиваются пчелиные матки и рабочие пчелы.
Наибольшую ценность матка представляет в первые
два года своей жизни. Начиная же с третьего года количество откладываемых ею
яиц быстро уменьшается. При этом старые матки откладывают много не
оплодотворённых яиц, из которых выводятся трутни. Такие матки называются
отрутневевшими. Пчелиные матки живут до пяти лет и более.
Рабочие пчёлы. Пчелиная семья состоит в основном
из рабочих пчёл. В хорошей семье зимой их насчитывается обычно 20-30 тыс., а
летом до 60-80 тыс. и больше. Все рабочие пчёлы в семье - сёстры, происшедшие
от одной матки. Они, как и пчелиная матка, являются женскими особями; однако, в
отличие от матки, половые органы у рабочих пчёл недоразвиты. Развивающихся из
яиц личинок выращивают рабочие пчёлы.
При внезапной гибели маток и отсутствии в гнезде
личинок предназначавшийся для них корм пчёлы-кормилицы потребляют сами, что
вызывает у них развитие яичников (каждый из 3-5, реже из 10-20 яйцевых
трубочек). Однако рабочие пчёлы не могут спариваться с трутнями. У них нет и
семяприёмника для сохранения сперматозоидов. Поэтому из отложенных такими
пчёлами неоплодотворённых яиц развиваются лишь трутни. Пчелы же с
функционирующими яичниками называются трутовками.
Рабочие пчёлы выполняют все работы внутри улья и
вне его. Они чистят гнездо, готовят ячейки сотов для откладки в них яиц маткой,
выделяют воск и строят новые соты, выкармливают личинок, поддерживают в улье
необходимую температуру, охраняют гнездо, собирают с цветков растений нектар и
пыльцу и приносят их в улей; Развитие рабочей пчелы со времени откладки яйца до
выхода взрослого насекомого длится в течение 21 суток, из них стадии: яйца - 3
суток, личинки в открытой ячейке - 6 суток, личинки и куколки в запечатанной
ячейке - 12 суток.
Трутни. Назначение трутней - осеменение молодых
маток, а также участие в тепловом балансе семьи. Развиваются они, как уже
отмечалось, из неоплодотворённых яиц, которые матка откладывает в ячейки сот,
имеющих несколько больший объём по сравнению с обычными пчелиными ячейками.
Личинки из яиц вылупляются к концу третьего дня. Пчёлы-кормилицы первые три дня
кормят личинку молочком, а затем - смесью мёда и перги. Стадия личинки в
открытой трутневой ячейке продолжается 6 суток, а стадия личинки и куколки в
запечатанной ячейке - 14 суток. Следовательно, развитие трутня совершается в
течение 24 суток.
Трутень заметно крупнее рабочей пчелы. Он весит
около 0,2 г. На выкармливание личинки трутня пчелы расходуют корма в три раза
больше, чем на вскармливание личинки рабочей пчелы. Поэтому на выращивание и
пропитание трутней семьи значительно увеличивают непроизводительный расход
кормов. Органы трутней не приспособлены для работы. У трутней короткий хоботок,
у них нет корзиночек для переноса пыльцы, нет восковых и других желез. У
трутней нет и жала, но они обладают сильными крыльями и очень большими сложными
глазами [Лебедев, Билаш, 1991].
Одиночные пчелы. По образу жизни и типам приспособления
к среде одиночные пчелы достигли широкого разнообразия и имеют большое значение
в природе и жизни человека. Термин «одиночные» пчелы свидетельствует о том, что
в отличие от медоносных пчел, ведущих общественный образ жизни, самка
самостоятельно строит гнездо с ячейками, куда собирает провизию для
выкармливания личинок. Забота о потомстве у одиночных пчел отсутствует
[Голиков, 2009].
Одиночные пчелы гнездятся чаще всего в почве,
хотя некоторые живут в толще древесины, в сухих стеблях тростника и т.п. В
типичном случае каждая самка сразу же после спаривания закапывается в почву,
прокладывая туннель диаметром чуть большим, чем диаметр ее собственного тела. В
зависимости от вида он уходит на глубину от 5-7 см до 1 м. Ячейки строятся либо
в ответвлениях туннеля, либо в его конце.
Завершив строительство ячейки, самка совершает
ряд вылетов к цветкам продолжительностью от нескольких минут до часа или более.
За каждый рейс она посещает множество цветков, собирая пыльцу, которую
переносит в гнездо на своих ногах или на нижней поверхности брюшка. Кроме того,
она обычно сосет и нектар, накапливаемый в зобу. Эти материалы помещаются в
ячейку и служат запасом пищи для личинки, которая будет здесь развиваться.
После того как накоплено необходимое количество провизии, пчела откладывает в
ячейку яйцо (как правило, на поверхность пищи), замуровывает вход в нее землей,
смешанной со слюной, и приступает к строительству следующей ячейки. Мать
никогда не увидит своего потомства; тем не менее перед нами прекрасный пример
заботы о нем. В зависимости от вида одиночная пчела за свою жизнь строит и
заполняет от 6 до 20 и более таких ячеек.
Представители семейства мегахилид (Megachilidae)
ведут себя несколько иначе. Пчелы этой группы обычно не выкапывают свою
собственную норку, а используют имеющиеся пустоты или строят гнездо на
поверхности. Самка заполняет ячейками найденное укрытие или прикрепляет их одну
к другой компактным комком. Это семейство включает в себя пчел-листорезов,
которые искусно сворачивают ячейки из кружочков, вырезаемых из листьев
определенных растений. В других отношениях поведение мегахилид примерно такое
же, как у прочих одиночных пчел.
После вылупления из яиц личинки питаются
запасенной провизией, которой обычно достаточно для роста и развития на
протяжении двух-трех недель. Затем личинка может окукливаться и превращаться в
имаго или же оставаться без изменений еще многие месяцы. Взрослая стадия многих
таких пчел длится всего несколько недель. Самка активна ранней весной, и уже к
концу сезона появляются зрелые личинки, которые не развиваются дальше на
протяжении всего лета и иногда даже осени и зимы. У видов с короткой личиночной
стадией имаго формируются к осени и зимуют в гнездах.
Многие одиночные пчелы устраивают изолированные
гнезда, находящиеся на значительном расстоянии от других. Однако некоторые виды
гнездятся скоплениями, хотя каждая самка занимается только своим гнездом.
Другим проявлением тенденции пчел к агрегации служит формирование спящих групп.
Самцы и - реже - самки многих одиночных видов собираются для ночевки в группы
(рои) на стеблях растений, обычно сцепляясь друг с другом челюстями. Таким
образом они могут ночевать на одном и том же месте много раз подряд [Пчелы,
2012].
Морфологические приспособления к опылению и
переносу пыльцы. На нижней части лба расположен наличник, под которым находится
ротовой аппарат, состоящий из пары нижних челюстей (максилл) и нижней губы,
нередко вытянутой в длинный хоботок, при помощи которого собирается нектар.
Тело большинства пчёл покрыто многочисленными
электростатическими ветвистыми ворсинками, способствующими прилипанию и
переносу пыльцы. Периодически они счищают с себя пыльцу, собирая их щёточками
(щетинковидными волосками, у большинства видов расположенными на лапках, а у
некоторых на брюшке) и затем переносят её в специальную корзиночку для пыльцы,
расположенную между задних лапок. подсолнечник
опыление цветение изолятор
Ноги у пчелиных служат не только для
передвижения и копания, а также для транспортировки пыльцы. У пчел, переносящих
пыльцу при помощи ног, последние обычно обильно опушены волосками, первый
членик задней лапки расширен, а широкая задняя голень часто с наружи углублена
и окаймлена волосками, образующими корзиночку. На внутренней стороне задней
голени имеется тибиальный гребень из шипиков, служащий для счесывания пыльцы в
корзиночку. У представителей рода Megachile
пыльца переносится на брюшной щеточке [Голиков, 2009]. Представители Colletidae
переносят пыльцу в зобике.
Суточная динамика лета в зависимости от
абиотических факторов и численности. Установлено, что численность пчелиных
широко варьирует в пределах суток, однако для подсолнечника всех сортов
отмечается совпадение периодов интенсивного посещения и спада. Активность
основной массы пчелиных достигает максимума к 10-12 ч, а затем, в наиболее
жаркие часы дня резко снижается. После 14-15 ч активность пчелиных
возобновляется, достигая второго более высокого максимума к 18 ч, причем шмели
и медоносная пчела летают до позднего вечера (20 ч), самыми последними
заканчивая летный день. Это можно хорошо пронаблюдать в таблице 2.
Наибольший же перепад суточной активности более
чем в шесть раз отмечается у одиночных пчел. Причина указанных межвидовых
различий заключается, очевидно, в разности экологических требований у разных
групп насекомых-опылителей.
Суточная динамика лета пчелиных на
подсолнечнике зависит от метеорологических условий. Так, при температуре 36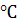 количество одиночных пчел на
учетной полосе не превышало двух особей. Максимальное количество опылителей (17
особей) зарегистрировано при температуре воздуха 23-27, которая оказалась наиболее
благоприятной для активного лета и работы пчел [Голиков, 2000].
количество одиночных пчел на
учетной полосе не превышало двух особей. Максимальное количество опылителей (17
особей) зарегистрировано при температуре воздуха 23-27, которая оказалась наиболее
благоприятной для активного лета и работы пчел [Голиков, 2000].
Таблица 2 - Средняя численность
опылителей подсолнечника в различное время суток [Голиков, 2008]
|
Время
суток, ч
|
Группа
опылителей, особей
|
|
Apis
mellifera
|
Bombus
sp.
|
одиночные
пчелы
|
|
8-10
|
8,5
(2)
|
6,6
(2)
|
3,2
(2)
|
|
13-15
|
4,0
(3)
|
3,4
(3)
|
0,8
(3)
|
|
18-20
|
11,0
(1)
|
7,2
(1)
|
5,1
(1)
|
|
Перепад
суточной активности, раз
|
2,8
|
2
|
6,4
|
Пчёлы могут переносить лишь
кратковременное воздействие высокой температуры. Длительное тепловое
воздействие (в течение более 30 минут при 46) вызывает необратимые изменения в
их организме, которые тем сильнее, чем длительнее период воздействия высоких
температур [Полевая работа пчел … , 2012].
Летная активность зависит от колебаний
влажности воздуха. На момент наблюдений с понижением ее (до 50%) численность
одиночных пчелиных также уменьшалась и составляла в среднем 2 особи на 100 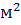 . С повышением относительной
влажности до 60% численность одиночных пчелиных возрастала до 7 особей/100 , а при повышении ее до 70%
численность опылителей увеличивалась и достигала 20 особей/100 . Вероятно, 70%-ная влажность
является оптимальной для полетов пчел (в период наблюдений температура воздуха
была 25-32) [Голиков, 2000].
. С повышением относительной
влажности до 60% численность одиночных пчелиных возрастала до 7 особей/100 , а при повышении ее до 70%
численность опылителей увеличивалась и достигала 20 особей/100 . Вероятно, 70%-ная влажность
является оптимальной для полетов пчел (в период наблюдений температура воздуха
была 25-32) [Голиков, 2000].
Также важно отметить, что летная
активность пчелиных значительно уменьшается перед выпадением осадков, и при их
выпадении она отсутствует.
Активность посещения подсолнечника
пчелиными зависит и от силы ветра: чем она выше, тем ниже активность пчелиных.
За весь период наблюдений наибольшая скорость ветра (10 м/с) наблюдалась лишь в
течение одного дня. В этот день насекомые не летали. Максимальное количество
опылителей наблюдалось при скорости ветра 1-2 м/с, при этом температура и влажность
воздуха находились в пределах от 25 до 29и от 55 до 65% соответственно
[Голиков, 2000].
1.4 Селекция подсолнечника
Подсолнечник - культура больших
возможностей. Современные сорта-популяции и гибриды при высокой агротехнике и
при благоприятных климатических условиях могут обеспечить урожай маслосемян
свыше 4,0 т/га с масличностью семян 53-54 %.
За многие десятилетия выращивания
подсолнечника он сам по себе изменился как по морфологическим признакам, так и
по биологическим особенностям. Современные сорта и гибриды имеют тонкую лузгу,
высокую масличность, толерантность к загущению, устойчивость к заразихе и
другим болезням.
Главной особенностью селекции
является использование закономерностей наследственности. Все улучшения
сортов-популяций и гибридов подсолнечника, которые удается получить
селекционеру, должны наследоваться и способствовать увеличению производства
растительного масла.
Современные сорта подсолнечника
являются сложными популяциями гетерозиготных растений, обладающими богатой
наследственностью. Они служат ценнейшим исходным материалом для получения
разнообразных самоопыленных линий. Для получения линий используют межлинейные
гибриды, обладающие комплексом ценных селекционных признаков, что дает
возможность получать линии, объединяющие лучшие признаки родительских форм
гибридов [Горьковая, 2011].
В селекции самоопыленных линий
предпочтение должно отдаваться автофертильным формам как наиболее пригодным при
развертывании первичного и промышленного семеноводства [Зайцев, 2009]. Такие
линии легче сохранять в процессе репродуцирования и целесообразно использовать
при селекции автофертильных гибридов.
Селекция подсолнечника бывает двух
направлений: создание сортов-популяций и гибридов.
Сорта-популяции - метод простого
индивидуального отбора с оценкой потомств и последующим направленным
переопылением, т.е. собственно селекция сортов-популяций по В.С.Пустовойту,
представляет собой по сути дела некое продолжение народной селекции, существовавшей
ранее. То есть в качестве исходного материала берется какая-то икс-популяция
или смесь популяций, представляющая собой чрезвычайно полиморфный,
гетерозиготный или, образно говоря, генетически «разношерстный» материал, из
которого чисто визуально, по каким-то субъективным признакам, импонирующим тому
или иному селекционеру, отбираются якобы лучшие растения или их корзинки и,
резервируя часть семян, высеваются на отдельном участке для дальнейшего их
сравнения. После такой оценки, из резервов лучших - отбираются еще более лучшие
и т.д., вплоть до объединения лучших семей, т.е. потомств с корзинок лучших
растений [О селекции сортов … , 2012].
Сорта-популяции подсолнечника цветут
свободно. Полезность и необходимость пчел, как опылителей, здесь неоспорима.
Это следует из вышесказанного.
Что же касается гибридов, исходным
материалом в их создании были лучшие сорта-популяции. Хотя уже давным-давно
последние не используются для этих целей, а настоящим исходным материалом
являются синтетические популяции или синтетики, полученные из наилучших
родительских линий и гибридов.
Исходная синтетическая популяция,
состоящая только из самых наилучших по всем признакам генотипов,
концентрирующая в себе максимальное количество полезных наследственных
факторов, преднамеренно расщепляется путем инбридинга, инцухта или самоопыления
в течение 5-7 поколений на самоопыленные линии с разбросом всех признаков
относительно исходной популяции, находящейся как бы в среднем состоянии, как в
отрицательную, так и положительную сторону. То есть объединенная и усредненная
масса генотипов в гетерозиготном состоянии как бы раскладывается в одну линию,
на весь спектр от положительных до отрицательных признаков, но уже в
гомозиготном константном состоянии. При этом в процессе самоопыления инбредные
линии подвергаются депрессии вплоть до летального исхода. И здесь как раз таки
задача селекционера взять в дальнейшую работу линии только положительной части
спектра.
Далее доведенные до константности,
т.е. способности сохранять при изоляции в неизменном виде накопленные признаки
десятилетиями линии, скрещивают между собой для выявления наивысшей степени
проявления эффекта гетерозиса и высокой комбинационной способности, не говоря
уже и обо всех хозяйственно и технологически полезных признаках. Таким образом,
создаются десятки тысяч линий с последующей выбраковкой худших из самых лучших
и производятся тысячи скрещиваний их между собой, с последующим испытанием и
выделением лишь десятков или максимум сотен гибридных комбинаций
предварительного испытания, и единицы или максимум десятки для конкурсного
испытания.
При селекции гибридов на участках
гибридизации есть мужские и женские линии. Женские представленные в двух
формах: А и Б. Вследствие скрещивания материнской и отцовской формы, образуется
межлинейный гибрид F1, который и используется в
производстве.
А-форма - растения с
цитоплазматической мужской стерильностью (ЦМС). Начиная с 1930-1932 гг., после
работ М.И. Хаджинова, М. Родса, случаи наследования признака мужской
стерильности описаны более чем у 80 видов покрытосеменных растений.
Использование мужской стерильности в практике обусловлено тем, что для
получения промышленных гибридов нет необходимости проводить кастрирование
цветков на материнских растениях.
У растений стерильной линии образуется
небольшое количество пыльцы, это позволяет провести генетический анализ. В
настоящее время ЦМС широко используется в сельскохозяйственной практике для
получения гибридных семян [Дубинин, 1986].
На участке размножения материнской
линии А-форму (ЦМС) скрещивают с Б-формой, которая является фертильным аналогом
(или закрепителем стерильности. Рекомендуется высаживать А-форму и Б-форму в
соотношении 8:6 соответственно. А-форма опыляется Б-формой, в результате чего
из семян А-формы опять получаются стерильные растения, а из Б-формы -
фертильные.
На участке гибридизации А-форма
(женская линия) скрещивается с Rf (мужская линия) -
восстановитель фертильности. В итоге А-форма дает семена для получения гибрида F1. Это есть
гетерозис - гибридная мощность, проявляющаяся в превосходстве гибрида над
обоими родительскими формами [Инге-Вечтомов, 1989]. На участке гибридизации
материнские и отцовские растения рекомендуется высаживать в соотношении 12:4
соответственно, так как отцовские растения ветвистые, и такое отношение
считается самым оптимальным.
Выделившийся по всем параметрам
гибрид необходимо теперь подготовить к промышленному семеноводству. То есть на
основе материнской линии с использованием доноров стерильности создается путем
беккроссов или насыщающих скрещиваний стерильный аналог или абсолютно
аналогичная линия, но без тычинок, что даст в дальнейшем возможность 100%-ной
гибридизации. В то же время по отцовской линии создается также аналог -
восстановитель фертильности, который в последствии на участках гибридизации
опылит на 100% стерильный аналог своей пыльцой, при полной пространственной
изоляции от любого другого подсолнечника, и на следующий год уже в
производственных посевах хозяйств восстановит фертильность, т.е. восстановятся
или появятся снова пыльники или тычинки на растениях гибридов 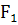 , выращенных из семян, убранных с
растений материнской стерильной линии.
, выращенных из семян, убранных с
растений материнской стерильной линии.
Таким образом, в отличие от сортов,
находящихся в состоянии постоянной изменчивости из-за расщепления и
последующего вырождения и деградации, гибриды можно поддерживать в неизменном
состоянии бесконечно долго до выведения лучших новых гибридов [О селекции
гибридов … , 2012].
В настоящее время сорт
рассматривается, как биологический способ производства, целостную генетическую
систему, способную к саморегуляции и самоорганизации в агрофитоценозе. Как
известно, компонентами гибридов подсолнечника являются самоопыленные линии,
полученные путем инцухта и их стерильные аналоги, созданные с помощью
беккроссных скрещиваний фертильной линии с ЦМС формой.
По продуктивности и другим полезным
признакам гибриды превышают сорта-популяции за счет гетерозиса при скрещивании
инцухт линий. Стратегия селекции гибридного подсолнечника базируется на создании
высокоурожайных самоопыленных линий с высокой устойчивостью к основным болезням
и паразитам [Кириченко, 2005].
2. Условия и методы исследований
.1 Природно-климатические
характеристики места исследования
Исследования проводили на центральной экспериментальной
базе ВНИИ масличных культур им. В.С. Пустовойта, г. Краснодар на 37-ом
пространственном изоляторе и в групповых сетчатых изоляторах в 2011 г.
Почва опытных полей института представлена
чернозёмом выщелоченным слабогумусным сверхмощным легкоглинистым. В пахотном
слое почвы содержится 3,3-3,5% гумуса. Почвы накапливают сравнительно небольшое
количество влаги, доступной для растений. Её запасы в двухметровом слое
составляют 694 мм, из них продуктивной влаги около 40%. Структура пахотного
слоя почвы - комковатая, в подпахотном горизонте - комковато-зернистая.
Весна ранняя, с возможностью заморозков до
начала мая. Лето продолжительное и жаркое. Осень обычно сухая в первой половине
и обильная осадками во второй. Тёплый и безморозный период продолжается до
декабря. Зима мягкая с неустойчивым снежным покровом и частыми оттепелями,
количество тёплых дней зимой достигает 65-70 дней .
Среднегодовая температура воздуха составляет
+11,4°С, средняя многолетняя температура самого холодного месяца (января) равна
-2,3ºС,
самого тёплого (июля) +23,4°С. В летние месяцы температура достигает 38-40ºС.
Продолжительность безморозного периода в условиях г. Краснодара достигает 193
дней (представлены данные метеостанции «Круглик»).
Главным абиотическим фактором, влияющим на
скорость развития подсолнечника, является температура. Для формирования урожая
наиболее благоприятна в период от цветения до созревания умеренная
среднесуточная температура 22-25˚С, которой обычно сопутствуют
благоприятные условия увлажнения. Данный район отвечает такому температурному
ограничению.
На рисунке 4 изображен график изменения
температур в год исследований на протяжении всего важного для вегетации
подсолнечника периода - с марта по август 2011 г. Благоприятная для прорастания
семян температура посевного слоя почвы составляет 10-12°С.
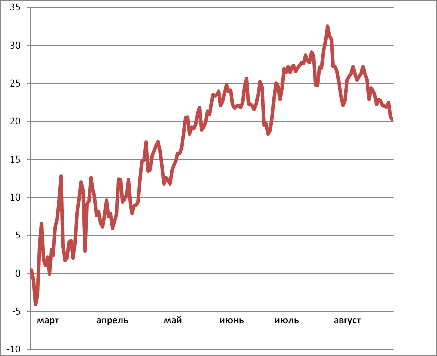
Рисунок 4 - График изменения среднесуточных
температур с марта по август 2011 г. в г. Краснодар
Температура воздуха летом в годы проведения
экспериментов была близка к среднемноголетним данным и составляла 23,7°С.
Средняя сумма осадков за год в зоне г.
Краснодара составляет 643 мм, в летние месяцы их выпадает около 180 мм. На
рисунке 5 изображено количество осадков, выпавших в период с марта по август
2011, по месяцам.
Центральная экспериментальная база ВНИИМК
расположена в центральной части Краснодарского края, относящейся к зоне
неустойчивого увлажнения. Поэтому здесь основным абиотическим фактором,
лимитирующим продуктивность подсолнечника, является дефицит влаги.
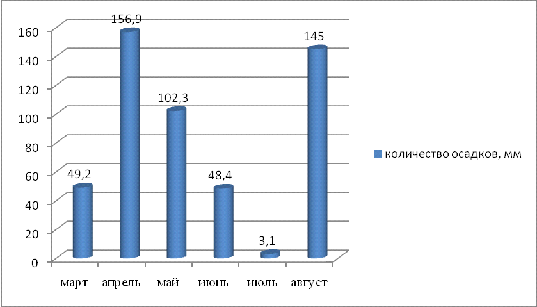
Рисунок 5 - Количество осадков, выпавших с марта
по август 2011г. в г. Краснодар
В период вегетации растений подсолнечника часто
наблюдаются засухи. Это можно пронаблюдать на рисунке 6, на котором
показывается количество и дни выпадения осадков в самый засушливый месяц
периода вегетации подсолнечника на 2011г. - июль.
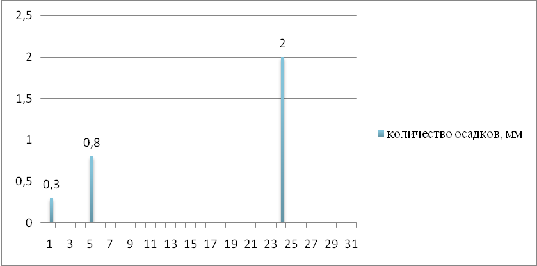
Рисунок 6 - Количество осадков, выпавших в июле
2011г. в г. Краснодар
Хотя подсолнечник считается засухоустойчивой
культурой в фазы вегетативного роста, однако он чувствителен к недостатку влаги
в период цветения и налива семян. При сильной засухе в период цветения большое
число заложившихся цветков, особенно в центральной части корзинки, не
расцветает. Следовательно, для наилучшего развития растений подсолнечника
желательно глубокое промачивание почвы в предпосевной период, умеренные осадки
в течение вегетационного периода до начала налива семян, а также отсутствие
осадков и низкая относительная влажность воздуха в конце периода налива семян.
2.2 Материалы и методы исследований
Пространственный изолятор. Учет опылителей
проводился 7, 11 и 13 июля 2011 г. на 37-ом пространственном изоляторе (рисунок
7), который является участком размножения Б-формы.
Здесь применялась пространственная изоляция при
производстве гибридных семян подсолнечника, когда семеноводческие посевы
размещают на определенном расстоянии, например, от товарных посевов в
соответствии с рекомендованными нормами пространственной изоляции. Эти нормы в
разных странах и их регионах разные. Причем каждая селекционно-семеноводческая
фирма имеет свои собственные нормы пространственной изоляции от зарослей
дикорастущего однолетнего подсолнечника.
Изоляция не всегда может быть обеспечена, в
результате этого получение чистого семенного материала превращается в
труднейшую задачу. Получение чистого семенного материала самоопыленных линий
подсолнечника - одно из основных условий конкурентоспособности гибридного
посевного материала [Новые российские патенты … , 2011].
Расстояние от 37-изолятора до ближайшего поля
подсолнечника (рисунок 8) - 1 км. Важно отметить, что между 37-ым
пространственным изолятором и ближайшим полем подсолнечника проходит трасса, по
которой активно движется транспорт, и две лесополосы, что является препятствием
для опылителей, которые могут принести на участок размножения гибрида
«чужеродную» пыльцу.
В последующем семена, полученные на 37-ом
пространственном изоляторе изучили на генетическую чистоту: определили состав
токоферолов 100 семянок. В результате проведенных опытов было выяснено, что все
100 семянок образовались от собственного опыления, т. е. были типичными для
линии. Следовательно, все правила пространственной изоляции были соблюдены.
Размеры 37-ого изолятора - 100 м на 30 м: 0,3
га. Это участок гибридизации линии ВК 876. Среднее количество растений на поле
- 8000. Условия: свободное цветение при пространственной изоляции.
Исследования проводились маршрутным методом,
основанным на методике Фасулати. Этот метод наиболее рационален. При этом
способе наблюдатель в течение 30 минут двигается по площадке и учитывает всех
замеченных опылителей [Фасулати, 1971].
Наша методика заключалась в следующем: через
поле проходят два человека - один ведет запись, второй диктует. Ведется учет
опылителей, сидящих в конкретный момент на корзинке подсолнечника. Записываются
данные 100 корзинок, которые находятся на втором-четвертом дне цветения, причем
растения выбираются с тем учетом того, чтобы пройти все поле.
Учитывается количество и состав опылителей, для
определения которые были собраны. До определения, для удобства, опылителям дали
обозначения, в соответствии с их морфологическими признаками. Также учитывались
время (10-11часов), облачность и температура воздуха (приложение А).
Сбор насекомых осуществлялся энтомологическим
сачком, который изготавливается самостоятельно и состоит из обруча, мешка и
ручки. Пойманную пчелу целесообразно извлекать пробиркой, так как ее укусы
болезненны. Пробирку с насекомым закрывают ваткой, смоченной в эфире, и
переворачивают вверх дном. Когда пчела достаточно обездвижена, ее помещают в
морилку. Морилка представляет собой небольшую стеклянную банку. Для поглощения
влаги на дно морилки помещается 2-3 кружочка фильтровальной бумаги по диаметру
дна, а внутрь - до 10 полосок сложенной мелкой гармошкой фильтровальной бумаги
размером 100 на 10 мм. Стеклянная емкость плотно прикрывается пробкой. К
внутренней стороне пробки прикрепляется небольшой комочек ватки и марли,
которые смачиваются эфиром [Голиков, 2009].
Групповой cетчатый
изолятор. Также исследования проводились в групповом сетчатом изоляторе,
который представляет собой металлическую конструкцию, обтянутую специальной
сеткой (рисунок 9). Размер исследуемого изолятора - 30 м на 5 м: 150 . Это участок гибридизации гибрида
Окси. Между собой скрещивается мать - ВК176 и отец - ВК195.
Предназначение сетчатого изолятора
заключается в том, чтобы не допустить попадание внутрь ни одного насекомого,
способного переносить пыльцу. Могут допускаться лишь те насекомые, которые
изначально были внутри сетчатого изолятора, так как они будут переносить пыльцу
с растений на растения внутри него самого, а не с растений, находящихся за его
приделами. Но это только теоретически. На практике же главные опылители
подсолнечника - пчелиные не живут в таких изоляторах, из-за другого
температурного режима и т. д. Из этого следует то, что должен вестись очень
строгий контроль за целостностью сетчатого изолятора.
Для опыления подсолнечника в
сетчатом изоляторе существует определенная порода пчел. Но так как это требует
больших экономических затрат и соблюдения строжайших правил их размещения и
«чистоты» опылителей. Такой способ применяется крайне редко.
На данном участке гибридизации
скрещивается мать - А-форма и отец - Rf
(восстановитель фертильности). Как можно видеть на рисунке 9, расположение
мужских и женских линий производится следующим образом по рядам: растения
женские, мужские, женские, женские, мужские, женские. Растений мужской линии
значительно меньше, так как они ветвистые, и такое соотношение считается
оптимальным.
В сетчатом изоляторе опыление
производится вручную путем либо сбора пыльцы с растений мужской линии и
нанесения ее на растения женской, либо при помощи наклона и соприкосновения
корзинки матери к корзинке отца. При входе человека в сетчатый изолятор должны
быть предусмотрены следующие правила: на одежде и руках не должно быть
«чужеродной» пыльцы; при открытии изолятора в него не должны попасть насекомые
из вне.
3. Особенности опыления
подсолнечника при свободном цветении и в условиях сетчатого изолятора
.1 Свободное цветение на
пространственно-изолированном участке
В результате исследований были
определены семейства собранных насекомых, которых на 37-ом пространственном
изоляторе, на момент исследований, было собрано 7 видов (в соответствии с
таблицей 4). Они представлены в таблице 3.
Таблица 3 - Представители насекомых,
собранных на 37-ом изоляторе
|
Отряд
перепончатокрылые (Hymenoptera)
|
Отряд
двукрылые (Diptera)
|
Отряд
чешуекрылые (Lepidoptera)
|
|
Семейство,
род, вид
|
|
сем.
Apoidae A. mellifera L.; Bombus terrestris L.
|
сем.
Syrphoidea Syrphus ribesi L.
|
сем.
Nymphalidae род Aglais
|
|
сем.
Colletidae род
Prosopis
|
-
|
-
|
|
сем.
Halictidae род
Halictidus
|
-
|
-
|
|
cем.
Andrenidae Andrena flavipes Pz.
|
-
|
-
|
При подсчетах и осреднении собранного материала
оказалось, что на долю медоносной пчелы (A.
mellifera) из всех
опылителей, присутствующих на 37-ом пространственном изоляторе, приходится 61%.
Следовательно остальных опылителей - 39%, в числе которых Prosopis
sp. - 7%, Halictidus
sp. - 27%, Aglais
- 4%, B. terrestris
L. - 0,3%, S.
ribesi - 0,3%, A.
flavipes - 0,3% (рисунок
10).
Из вышесказанного видно, что главным опылителем
подсолнечника является медоносная пчела (A.
melliferа). Далее по
значимости для опыления подсолнечника следуют одиночные пчелы (Prosopis
sp., Halictidus
sp., A.
flavipes). После одиночных
пчел конкретно на 37-ом пространственном изоляторе следуют B.
terrestris; Aglais
sp. и S.
ribesi.
Ведение учета опылителей - их количество и
представители семейств, показаны в таблице 4:
Таблица 4 - Учет опылителей на 37-ом изоляторе
7 июля 2011 г. 11 июля 2011 г. 13
июля 2011 г.
ясно ясно
ясно (ветрено)
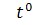 27-28°С
27-28°С 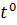 28°С 28°С
28°С 28°С
|
№
|
1
|
2
|
3
|
4
|
5
|
6
|
7
|
|
№
|
1
|
2
|
3
|
4
|
5
|
6
|
7
|
|
№
|
1
|
2
|
3
|
4
|
5
|
6
|
7
|
|
1
|
|
1
|
|
|
|
|
|
|
1
|
1
|
|
|
|
|
|
|
|
1
|
|
|
|
|
|
|
|
|
2
|
|
|
|
|
|
|
|
|
2
|
|
|
1
|
|
|
|
|
|
2
|
|
|
|
|
|
|
|
|
3
|
2
|
|
|
|
|
|
|
|
3
|
|
1
|
|
|
|
|
|
|
3
|
1
|
|
|
|
|
|
|
|
4
|
|
|
1
|
|
|
|
|
4
|
|
|
|
|
|
|
|
|
4
|
|
|
|
|
|
|
|
|
5
|
1
|
|
|
|
|
|
|
|
5
|
|
|
|
|
|
|
|
|
5
|
|
|
|
|
|
|
|
|
6
|
|
|
|
|
|
|
|
|
6
|
|
1
|
|
|
|
|
|
|
6
|
|
1
|
|
|
|
|
|
|
7
|
|
1
|
|
|
|
|
|
|
7
|
2
|
|
|
|
|
|
|
|
7
|
|
|
|
|
|
|
|
|
8
|
1
|
|
|
|
|
|
|
|
8
|
1
|
|
|
|
|
|
|
|
8
|
1
|
|
|
|
|
|
|
|
9
|
1
|
|
|
|
|
|
|
|
9
|
|
|
|
|
|
|
|
|
9
|
|
|
|
|
|
|
|
|
10
|
1
|
|
|
|
|
|
|
|
10
|
1
|
|
|
|
|
|
|
|
10
|
1
|
|
|
|
|
|
|
|
11
|
|
|
1
|
|
|
|
|
|
11
|
|
1
|
|
|
|
|
|
|
11
|
|
1
|
|
|
|
|
|
|
12
|
|
|
|
|
|
|
|
|
12
|
|
|
|
|
|
|
|
|
12
|
|
|
|
|
|
|
|
|
13
|
1
|
|
|
|
|
|
|
|
13
|
|
|
1
|
|
|
|
|
|
13
|
1
|
|
|
|
|
|
|
|
14
|
|
|
1
|
|
|
|
|
|
14
|
1
|
|
|
|
|
|
|
|
14
|
1
|
|
|
|
|
|
|
|
15
|
1
|
|
|
|
|
|
|
|
15
|
|
|
1
|
|
|
|
|
|
15
|
|
|
1
|
|
|
|
|
|
16
|
1
|
|
1
|
|
|
|
|
|
16
|
|
|
|
|
|
|
|
|
16
|
|
|
|
|
|
|
|
|
17
|
|
|
|
1
|
|
|
|
|
17
|
|
|
|
|
|
|
|
|
17
|
1
|
|
|
|
|
|
|
|
18
|
1
|
|
|
|
|
|
|
|
18
|
1
|
|
|
|
|
|
|
|
18
|
1
|
|
|
|
|
|
|
19
|
|
|
|
|
|
|
|
|
19
|
1
|
|
|
|
|
|
|
|
19
|
|
|
|
|
|
|
|
|
20
|
1
|
|
1
|
|
|
|
|
|
20
|
|
|
|
|
|
|
|
|
20
|
1
|
|
|
|
|
|
|
|
21
|
1
|
|
|
|
|
|
|
|
21
|
|
|
1
|
|
|
|
|
|
21
|
|
|
|
|
|
|
|
|
22
|
|
|
1
|
|
|
|
|
|
22
|
1
|
1
|
|
|
|
|
|
|
22
|
|
|
2
|
|
|
|
|
|
23
|
|
|
|
|
|
|
|
|
23
|
|
|
1
|
|
|
|
|
|
23
|
1
|
|
|
|
|
|
|
|
24
|
1
|
|
|
|
|
|
|
|
24
|
|
|
|
|
|
|
|
|
24
|
|
|
|
|
|
|
|
|
25
|
|
|
|
|
|
|
|
|
25
|
|
|
|
|
|
|
|
|
25
|
|
1
|
|
|
|
|
|
|
26
|
|
|
|
|
|
|
|
|
26
|
1
|
|
|
|
|
|
|
|
26
|
1
|
|
|
|
|
|
|
|
27
|
|
|
|
|
|
|
|
|
27
|
|
|
|
|
|
|
|
|
27
|
|
|
1
|
|
|
|
|
|
28
|
1
|
|
|
|
|
|
|
|
28
|
|
|
|
|
|
|
|
|
28
|
1
|
|
|
|
|
|
|
|
29
|
|
|
|
|
|
|
|
|
29
|
|
|
|
|
|
|
|
|
29
|
1
|
|
|
|
|
|
|
|
30
|
|
|
|
|
|
|
|
|
30
|
1
|
|
|
|
|
|
|
|
30
|
|
|
|
|
|
|
|
|
31
|
|
|
|
|
|
|
|
|
31
|
1
|
|
1
|
|
|
|
|
|
31
|
2
|
|
|
|
|
|
|
|
32
|
1
|
|
|
|
|
|
|
|
32
|
|
|
|
|
|
|
|
|
32
|
|
|
|
|
|
|
|
|
33
|
|
|
1
|
|
|
|
|
|
33
|
1
|
|
|
|
|
|
|
33
|
1
|
|
|
|
|
|
|
|
34
|
1
|
|
|
|
|
|
|
|
34
|
|
|
|
|
|
|
|
|
34
|
|
|
|
|
|
|
|
|
35
|
1
|
|
|
|
|
|
|
|
35
|
|
|
|
|
|
|
|
|
35
|
1
|
|
|
|
|
|
|
|
36
|
|
1
|
|
|
|
|
|
|
36
|
|
|
|
|
|
|
|
|
36
|
1
|
|
|
|
|
|
|
|
37
|
|
1
|
|
|
|
|
|
|
37
|
|
|
|
|
|
|
|
|
37
|
|
|
|
|
|
|
|
|
38
|
1
|
|
|
|
|
|
|
|
38
|
1
|
|
|
|
|
|
|
|
38
|
1
|
|
|
|
|
|
|
|
39
|
1
|
|
|
|
|
|
|
|
39
|
1
|
|
|
|
|
|
|
|
39
|
|
|
|
|
|
|
|
|
40
|
|
|
|
|
|
|
|
|
40
|
|
|
1
|
|
|
|
|
|
40
|
1
|
|
|
|
|
|
|
|
41
|
1
|
|
1
|
|
|
|
|
|
41
|
1
|
|
|
|
|
|
|
|
41
|
|
|
|
|
|
|
|
|
42
|
1
|
|
|
|
|
|
|
|
42
|
|
|
|
|
|
|
|
|
42
|
1
|
|
|
|
|
|
|
|
43
|
|
|
1
|
|
|
|
|
|
43
|
|
|
|
|
|
|
|
|
43
|
|
|
|
|
|
|
|
|
44
|
|
|
|
|
|
|
|
|
44
|
1
|
|
|
|
|
|
|
|
44
|
1
|
|
|
|
|
|
|
|
45
|
|
|
|
|
|
|
|
|
45
|
|
|
|
|
|
|
|
|
45
|
|
|
|
|
|
|
|
|
46
|
|
|
|
|
|
|
|
|
46
|
|
|
1
|
|
|
|
|
|
46
|
1
|
|
|
|
|
|
|
|
47
|
|
|
|
|
|
|
|
|
47
|
|
1
|
1
|
|
|
|
|
|
47
|
1
|
|
|
|
|
|
|
|
48
|
1
|
|
|
|
|
|
|
48
|
1
|
|
1
|
|
|
|
|
|
48
|
2
|
|
|
|
|
|
|
|
49
|
1
|
|
|
|
|
|
|
|
49
|
|
|
|
|
|
|
|
|
49
|
|
|
1
|
|
|
|
|
|
50
|
1
|
|
|
|
|
|
|
|
50
|
1
|
|
|
|
|
|
|
|
50
|
|
|
|
|
|
|
|
|
51
|
|
|
1
|
|
|
|
|
|
51
|
|
|
|
|
|
|
1
|
|
51
|
1
|
|
|
|
|
|
|
|
52
|
1
|
|
|
|
|
|
|
|
52
|
|
|
|
|
|
|
|
|
52
|
|
|
|
|
|
|
|
|
53
|
1
|
|
|
|
|
|
|
|
53
|
|
|
|
|
|
|
|
|
53
|
|
|
1
|
|
|
|
|
|
54
|
|
|
|
1
|
|
|
|
|
54
|
|
|
1
|
|
|
|
|
|
54
|
2
|
|
|
|
|
|
|
|
55
|
1
|
|
|
|
|
|
|
|
55
|
|
|
|
|
|
|
|
|
55
|
|
|
|
|
|
|
|
|
56
|
|
|
1
|
|
|
|
|
|
56
|
|
|
|
|
|
|
|
|
56
|
1
|
|
1
|
|
|
|
|
|
57
|
|
|
|
|
|
|
|
|
57
|
|
|
|
|
|
|
|
|
57
|
|
|
|
|
|
|
|
|
58
|
|
|
|
|
|
|
|
|
58
|
|
|
|
|
|
|
|
|
58
|
|
|
|
|
|
|
|
|
59
|
1
|
|
|
|
|
|
|
|
59
|
|
|
|
|
|
|
|
|
59
|
|
|
|
|
|
|
|
|
60
|
|
|
|
|
|
|
|
|
60
|
1
|
|
|
|
|
|
|
|
60
|
|
|
|
|
|
|
|
|
61
|
1
|
|
|
|
|
|
|
|
61
|
|
|
|
|
|
|
|
|
61
|
1
|
|
|
|
|
|
|
|
62
|
|
|
|
|
|
|
|
|
62
|
1
|
|
|
|
|
|
|
|
62
|
|
1
|
|
|
|
|
|
63
|
|
|
|
|
|
|
|
|
63
|
|
|
|
|
|
|
|
|
63
|
|
|
1
|
|
|
|
|
|
64
|
|
|
|
|
|
|
|
|
64
|
1
|
|
|
|
|
|
|
|
64
|
1
|
|
|
|
|
|
|
|
65
|
1
|
|
|
|
|
|
|
|
65
|
|
|
2
|
|
|
|
|
|
65
|
1
|
|
|
|
|
|
|
|
66
|
1
|
|
|
|
|
|
|
|
66
|
|
|
|
|
|
|
|
|
66
|
1
|
|
1
|
|
|
|
|
|
67
|
|
|
|
|
|
|
|
|
67
|
|
|
1
|
|
|
|
|
|
67
|
1
|
|
|
|
|
|
|
|
68
|
1
|
|
|
|
|
|
|
|
68
|
1
|
|
|
|
|
|
|
|
68
|
|
|
|
|
|
|
|
|
69
|
|
|
1
|
|
|
|
|
|
69
|
|
|
|
|
|
|
|
|
69
|
1
|
|
|
|
|
|
|
|
70
|
|
|
|
|
|
|
|
|
70
|
|
|
|
|
|
|
|
|
70
|
|
|
|
|
|
|
|
|
71
|
|
|
|
|
|
|
|
|
71
|
|
|
1
|
|
|
|
|
|
71
|
|
|
1
|
|
|
|
|
|
72
|
|
|
|
|
|
|
|
|
72
|
1
|
|
|
|
|
|
|
|
72
|
|
|
|
|
|
|
|
|
73
|
1
|
|
|
|
|
|
|
|
73
|
|
|
|
|
|
|
|
|
73
|
1
|
|
|
|
|
|
|
|
74
|
|
|
|
1
|
|
|
|
|
74
|
|
|
|
|
|
|
|
|
74
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
Примечание: № - номер корзинки;1 - Apis
mellifera
L.; 2 - Prosopis
sp.; 3 - Halictidus
sp.; 4 - Aglais
sp.; 5 - Bombus
terrestris
L; 6 - Syrphus
ribesi L.;
7 - Andrena
flavipes
Pz.
Рисунок 11 хорошо иллюстрирует отличие
медоносной пчелы от одиночных. Медоносная пчела в основном питается нектаром,
следствием чего является крупный хоботок. Одиночные пчелы питаются в основном
пыльцой, для переноса которой у них имеются специальные корзиночки. Нектар
нужен им для создания «хлебца», соотношение пыльцы и нектара которого варьирует
у разных видов.
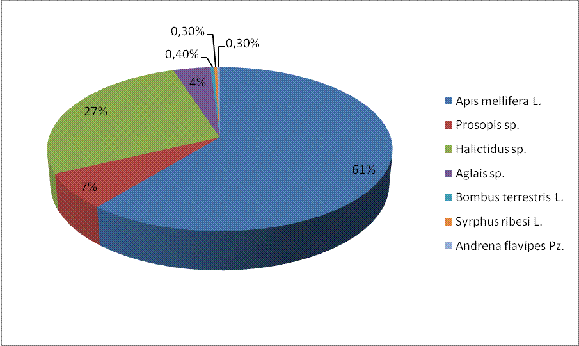
Рисунок 10 - Соотношение опылителей на 37-ом
изоляторе
3.2 Цветение под групповыми
сетчатыми изоляторами
В групповом сетчатом изоляторе опыление
подсолнечника производилось вручную. Это связано прежде всего с неблагоприятным
микроклиматом, из-за которого пчелы в нем существовать не могут, так как
главным лимитирующим фактором их опылительной активности является температура,
которая в свою очередь в групповом сетчатом изоляторе выше. Ветер как опылитель
здесь тоже играет очень небольшую роль вследствие наличия сетки.
Следовательно, без человеческого участия в
опылении подсолнечника, находящегося в групповом сетчатом изоляторе, не
обойтись (рисунок 12). С помощью такого опыления выполненость семян в корзинке
удалось увеличить до 70-75%, что приближено к значению этого показателя при
свободном цветении (80-85%). Без участия человека в опылении подсолнечника,
находящегося в сетчатом изоляторе, этот показатель был бы приближен к 1.
Опыление в сетчатом изоляторе проводилось через
день. Также велся строжайший контроль за целостностью изолятора, во избежание
попадания «чужеродной» пыльцы.
Впоследствии опыления вручную подсолнечника,
цветущего в групповом сетчатом изоляторе, выполненость семян в корзинках
оказалась хорошей - примерно 70%. Как показал определенный на 100 семян состав
токоферолов, «чужеродной» пыльцы обнаружено не было. Это говорит о том, что все
правила размножения гибрида при помощи сетчатого изолятора были соблюдены.
Заключение
В данной работе были рассмотрены особенности
опыления подсолнечника при свободном цветении и в условиях группового сетчатого
изолятора. Эти вопросы важны в селекции подсолнечника, основной целью которой
является улучшение сортов-популяций и гибридов подсолнечника для увеличения
производства растительного масла.
Селекция подсолнечника бывает двух направлений:
создание сортов-популяций и гибридов. При свободном цветении подсолнечника,
будь это выращивание сортов-популяций или гибрида, решающую роль в опылении
играют пчелиные, наиболее многочисленными из которых является медоносная пчела
(Apis mellifera
L.), процент которой
на исследуемом поле составил 61% от всех опылителей.
Также большой вклад в опыление подсолнечника
вносят одиночные пчелы, процент которых на исследуемом поле составил 35 % от
всех опылителей. Но их небольшое по сравнению с медоносной пчелой количество
объясняется ограничением дальности их полетов (примерно 200-300 м).
Для получения большего количества семян,
обладающих высокой генетической чистотой, в селекции используют групповые
сетчатые изоляторы. Они предотвращают попадание пыльцы с других растений,
переносимой насекомыми - прежде всего пчелами, а также ветром.
Относительно опыления в условиях сетчатого
изолятора, все иначе. Пчелиные там обитать не могут из-за неблагоприятного
микроклимата в изоляторе, в основном - высокой температуры. Есть только
определенные породы, которые способны обитать в этих условиях, но это требует
немалых экономических затрат и соблюдения строгих правил. Поэтому опыление
происходит при помощи человека путем сбора пыльцы с растений мужских линий и
нанесения ее на рыльца материнских растений. Конечно, это требует определенных
трудозатрат, но этот способ является самым оптимальным.
Так как сетчатый изолятор является участком
гибридизации, необходимо следить за целостностью изолятора и исключать
попадание в него насекомых из вне. Также необходимо исключить попадание в
изолятор «чужеродной» пыльцы с человеком, входящим в изолятор.
Список используемых источников
Арнольдова О.Н. К биологии цветения
подсолнечника в связи с техникой его скрещивания // Журнал опытной агрономии
Юго-Востока. 1982. Т. 2. Вып. 1. С. 18.
Богоявленский С.Г. Пчелы и
подсолнух. Записки Воронежского с.-х. института. Воронеж, 1935. Т. 1. 237 с.
Володарский Н.И, Газенко А.И. О
стадийной неоднородности тканей у подсолнечника // Ботан. журн. 1960. Т. 45.
№5. С. 743.
Гаврилова В.А. Анисимова И.Н.
Генетика культурных растений. Подсолнечник. СПб., 2003. 209 с.
Голиков В.И. Экологические
особенности опыления подсолнечника пчелиными // Масличные культуры.
Научно-технический бюллетень Всероссийского научно-исследовательского института
масличных культур. Краснодар, 2008. Вып. № 2. С. 27-29.
Голиков В.И. Фауна Кубани: одиночные
пчелы. Краснодар, 2009. 192 с.
Голиков В.И. Экологические основы
опыления некоторых полевых и плодовых культур пчелиными в Западном
Предкавказье. Краснодар, 2000. 192 с.
Горьковая Е.Г. Влияние
автофертильности исходного материала на завязываемость в первых поколениях
инбредных линий // Сборник материалов 6-ой международной конференции молодых
ученых и специалистов. Краснодар, 2011. С. 61-64.
Дубинин Н.П. Общая генетика. М.,
1986. 560 с.
Дьяков А.Б. Физиология
подсолнечника. Краснодар, 2004. 76 с.
Зайцев А.Н. Перспективный исходный
материал для селекции гибридов подсолнечника на автофертильность и
пчелопосещаемость // Масличные культуры. Научно-технический бюллетень ГНУ ВНИИ
масличных культур. Краснодар, 2009. №1 (140). С. 5.
Инге-Вечтомов С.Г. Генетика с
основами селекции. М., 1989. 591 с.
Кириченко В.В. Селекция и
семеноводство подсолнечника (Helianthus
annuus L.).
Харьков, 2005. 385 с.
Кривошлыков К.М. Анализ сложившегося
состояния и перспектив производства масличного сырья в Российской Федерации //
Сб. докладов 3-й международной конференции молодых ученых и специалистов
«Актуальные вопросы селекции, технологии и переработки масличных культур».
Краснодар, 2005.119 с.
Лебедев В.И., Билаш Н.Г. Биология
медоносной пчелы. М., 1991. 239 с.
Малюга Н.Г., Квашин А.А., Загорулько
А.В. Подсолнечник. Биология и агротехника выращивания на юге России:
монография. Краснодар, 2011. 302 с.
Масличные
культуры. Подсолнечник [Электронный ресурс]. URL: <http://www.meedov.ru/plantnectar/field/1.html> (дата
обращения 6.06.12).
Научный
отчет ВНИИМК 1934-1935 гг. по группе подсолнечника сектора селекции. Краснодар,
1936. 169 с.
Никитчин
Д.И. Подсолнечник. Биохимия, селекция, возделывание. Пологи, 2002. 494 с.
Новые
российские патенты (полные тексты). Способ получения генетически чистых семян
материнской формы для гибрида подсолнечника [Электронный ресурс]. URL:
http://partkom.com/patent/ru2370948/ (дата обращения 19.06.12).
О
селекции гибридов и их преимуществах. Селекционно-семенноводческая фирма
«Российская гибридная индустрия» [Электронный ресурс]. URL:
<http://rhifinal.narod.ru/preim.htm> (дата обращения 5.06.12).
О
селекции сортов и их недостатках. Селекционно-семенноводческая фирма
«Российская гибридная индустрия» [Электронный ресурс]. URL:
<http://rhifinal.narod.ru/nedost.htm> (дата обращения 5.06.12).
Определитель
насекомых Европейской части СССР. Т. 3 (1) / под ред. Медведева Г.С. СПб.,
1978. 584 с.
Плачек
Е.М. Формообразовательные процессы у подсолнечника под влиянием гибридизации и
инцухта // Тр. всес. съезда по генетике, селекции, семеноводству и
животноводству. М., 1930. Т. 2. С. 395.
Погода
и климат // Погода в Краснодаре [Электронный ресурс]. URL:
<http://www.pogoda.ru.net/monitor.php?id=34927&month=3&year=2011>
(дата обращения 17.06.12).
Полевая
работа пчел // Сайт воронежского пчеловодства [Электронный ресурс]. URL:
<http://medovyi.spas9.ru/s_18.htm> (дата обращения 18.06.12).
Подсолнечник.
Достижения селекции // Селекция полевых культур. Создание высокоурожайных
сортов [Электронный ресурс]. URL: <http://selekcija.ru/podsolnechnik-dostizheniya-selekcii.html>
(дата обращения 17.06.12).
Подсолнечник
/ под ред. В.С. Пустовойт. М., 1975. 592 с.
Подсолнечник
/ под ред. И.А Минкевич, Т.Т. Демиденко, В.С. Пустовойт. Краснодар, 1940. 310
с.
Поправке
С. А. Растения и пчелы. М., 1985. 240 с.
Постарнак
Ю.А. Правила выполнения и оформления выпускной квалификационной работы.
Краснодар, 2009. 104 с.
Пустовойт
В.С. Масличный подсолнечник. М., 1928. 305 с.
Пустовойт
В.С. Руководство по селекции и семеноводству масличных культур. М., 1967. 235
с.
Пчелы
// Энциклопедия Кругосвет [Электронный ресурс]. URL:
http://www.krugosvet.ru/node/38410?page=0,2#part-7
<http://www.krugosvet.ru/node/38410?page=0,2> (дата обращения 15.06.12).
Радченко
В.Г. Биология пчел (Hymenoptera, Apoidea). СПб,
1994. 350 с.
Словарь
генетических терминов. М., 2007. 508 с.
Строение
рабочей пчелы [Электронный ресурс]. URL: <http://www.krugosvet.ru/images/1000175_1560_002.gif> (дата
обращения 18.06.12).
Фасулати
К.К. Полевое изучение наземных беспозвоночных. М., 1971. 424 с.
Фрувирт
К. Подсолнечник (H. annuus L.). Селекция
кукурузы, кормовой свеклы и др. корнеплодов, масличных растений и кормовых
злаков. СПб., 1914. 270 с.