|
Категория
|
2004
г.
|
2005
г.
|
Проект
|
|
Выростные
|
4,8
|
5,3
|
8,4
|
|
Нагульные
|
6,8
|
7,5
|
11,9
|
Глава III. Характеристика кутума (результаты
исследований)
.1 Морфология кутума

Рис 2. Общий вид кутума
Кутум - проходная рыба. Большую часть жизни
проводит в море и только для икрометания идет в реки, впадающие в Астрабадский
и Пехлевийский заливы, в реки Ленкоранку, Куру, Кейранчай, Самур, Сулак, Терек
и др. После создания водохранилища на Сулаке и постройки Чиркейской ГЭС, что
привело к значительному уменьшению твердого стока реки и другим изменениям ее
гидрологического режима, Сулак, куда прежде кутум не заходил, приобрел функции
важного нерестового водоема. В Сулаке кутум поднимается почти на 100 км и
размножается на галечных россыпях в районе Султангиюрт-Нечаевка вплоть до
плотины Чирюртовской ГЭС.
В Среднем Каспии кутум для нереста заходит также
в Дивичинский лиман и Самурский НВВ. После мелиорации Малого Кызылагачского
залива (1955 г.) р. Кумбашинка, являвшаяся ранее главным нерестилищем кутума в
юго-западной части Каспия, утратила свое значение. В настоящее время нерест
кутума происходит в сбросном канале, а часть производителей через этот канал
заходит в Кировское НВХ и нерестится в его прудах.
Кутум начинает входить в Малый Кызылагачский
залив с конца января, интенсивный ход наблюдается обычно в марте при
температуре воды 12,3-13,8°С, в конце апреля-начале мая нерестовый ход
заканчивается. Общая его продолжительность в различные годы составляла от 55 до
70 дней. В Самурский НВВ нерестовая миграция кутума начинается в конце
февраля-начале марта при температуре воды в море 2,5-3"С, достигает
максимума в третьей декаде марта- начале апреля при 4,5-8,5°С и заканчивается
во второй половине апреля при прогреве воды до 9-11°С.
Кутум, населяющий Каспий, морфометрически
впервые исследован в 1932 г. А.В. Морозовым (1959), который собрал материал во
время нерестового хода в р. Кумбашинка, а в 1953 г. исследован из уловов р.
Куры А.Н. Смирновым.
В марте 1963 г. проведены первые значительные
морфометрические измерения 100 экз. кутума в района Аграхана. Их меристические
признаки: D III 9-10, AIII 9-11, число чешуи в боковой линии 51-63, над боковой
линией 8-10, под боковой линией 4-6, число позвонков 40-41. По этим признакам
достоверных различий между самками и самцами нет. Однако различия
обнаруживаются при сравнении пластических признаков (табл. 6). У самок
каспийского кутума несколько больше высота тела, антеанальное расстояние,
длиннее грудной плавник, расстояние между грудным и брюшным, а также брюшным и
анальным плавниками; у самцов - длиннее голова, хвостовой стебель, больше
диаметр глаза, высота спинного плавника, длина анального и брюшного плавников,
а также нижняя лопасть йостового плавника-Кроме того, ко времени нереста тело
самцов сплошь покрывается острыми шипиками, чем они легко отличаются от самок,
после нереста шипики опадают в виде пленки.
Таблица 6. Морфологические признаки кутума
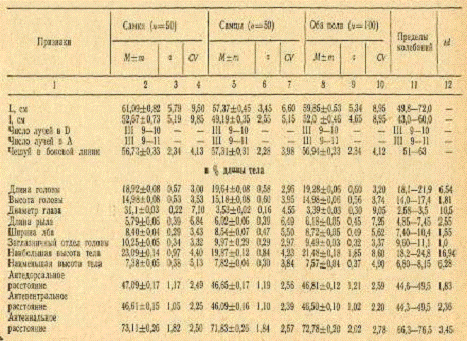
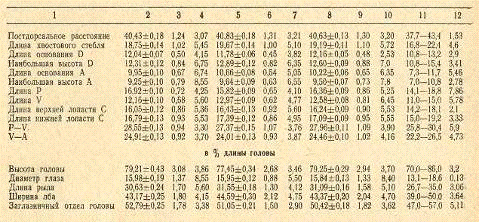
Вариабильность морфологических признаков у самок
из залива несколько выше, чем у самцов. Так, из 24 признаков у самок, судя по
коэффициенту CV, более вариабильными оказались 15 (62,6%), а у самцов - только
9 (37,4%). Средний коэффициент изменчивости признаков у самок составил 4,61, у
самцов - 4,30, эта разница достоверна при уровне значимости 0,05. Еще более
четко выражена такая зависимость у популяции кутума с западной прибрежной части
Среднего Каспия: у самок из 23 признаков более вариабельными оказались 19
(82,7%), а у самцов - только 4 (17,3%). Средний коэффициент изменчивости
признаков у самок среднекаспийского кутума составил 5,06% против 4,36% у
самцов.
Кутум населяет только Каспийское море. Ареал
приурочен к южным и западным берегам - от устья р. Терек на севере до залива
Горган на юге, основные скопления в - юго-западной части моря, прилегающей к
заливам Кирова и Пехлевийскому. Зона моря вдоль западного побережья Среднего и
Южного Каспия является постоянным местом обитания кутума в течение года,
встречается он здесь на глубине от 9 до 24 м. У восточного побережья Каспия в
небольшом количестве постоянно обитает в районах Красноводска, Карши, АИМ, на
приатрекском участке, в иранских водах кутум - основная промысловая рыба. В
середине апреля в очень небольшом количестве входит к дельту Волги, иногда и
выше, заходит в Каму, единичные экземпляры встречаются в апреле в устье Урала
(Берг, 1949). Акклиматизирован в водоемах Краснодарского края (Кубанские
лиманы), в Азовском море, на Украине (Дорошин, 1959; Жукинский, Балан , 1959;
Ужва, 1965).
.2 Экология нереста кутума
С 1901т., по сведениям А.Н. Державина (1949),
уловы кутума в районе Терека и Сулака достигали значительных величин. Так, в
1908- 1911 гг. только в Аграханском заливе и в бассейнах рек Терека, и Сулака
кутума отлавливали от 6,0 до 10 тыс. ц в, год. Начиная с 50-х годов, уловы
кутума в дагестанских водах уже не исчислялись тысячами центнеров и в настоящее
время (начиная с 1960- 1965 гг.) он по сути перестал в наших водах считаться
промысловой рыбой и учет ведется в общем улове частиковой рыбы. Кутум для
нереста в Нижне-Терские озера, по наблюдениям. Д.3. Демина (1946), в Самурское
озеро - Г.М. Магомедова (1970), заходит исключительно в ранне-весенний период,
причем ход начинается с середины марта и, постепенно возрастая, достигает
наибольшей интенсивности со второй половины апреля до наступления мая.
Кутум заходит в Терек в середине марта, в Волгу
в середине апреля, нерестится в феврале-мае, икру выметывает на заросли камыша
в разливах рек и в озерах. Как и вырезуб, кутум созревает на 4-5-м году жизни.
Плодовитость 27-280, в среднем 110 тыс. икринок. Диаметр зрелой икры 1,2-2,5
мм. Нерест одноразовый, при температуре воды 13-15о С. Икра откладывается на
растительность на глубинах до 0,8 м на мелководжьях заросших участков с
медленным течением. Мальки осенью постепенно скатываются в море. Подрастающие
рыбы, готовые к нересту, потом подходят к берегам. В море кутум питается
преимущественно моллюсками и крабами. (Промысловые рыбы СССР, 1949; Казанчеев,
1981).Нерестовый ход начинается при температуре воды 6,0- 7,2°, разгар хода -
при 12,3-13,8°. Общая продолжительность нерестового хода в различные годы -
55-70 дней. Икрометание происходит при температуре воды - 9-12°. Длительность
инкубации икры при температуре воды 5,5-24,8° составляет от 3 до 16 суток. По
данным Ю.А. Абдурахманова, инкубация икры кутума при температуре воды 8-11°
длится 11-15 дней, при 15-17° личинки выклевываются на 7-8-е сутки. Молодь
скатывается 14-20 дней, после рассасывания желточного мешка. Первое время
молодь держится в прибрежной зоне, затем отходит в море. Плодовитость рыб из
залива размером от 49 до 62 см составляет в среднем 101,3 тыс. икринок с
колебаниями от 90,8 до 119 тыс. По данным Ю. А. Абдурахманова (1962),
абсолютная плодовитость в зависимости от размера рыб колеблется от 27 до 280
тыс. икринок, в среднем 109,6 тыс. По материалам, собранным в 1973 г.,
установлено, что с увеличением длины, массы и возраста рыб абсолютная
плодовитость увеличивается, т. е. между плодовитостью и длиной, массой и
возрастом рыб существует прямая зависимость (табл. 7) Наименьшую плодовитость
(28500 икринок) имела рыба в возрасте 3 лет длиной 39 см, массой 810 г,
наибольшую (245 025) - соответственно 6 лет, 54 см, 2540 г.
После нереста рыбы по сбросному каналу
скатываются в море, в первые дни. нагуливаются в районе севера Аграхана, затем
уходят в море на глубину более 10 м.
Таблица 7.Плодовитость кутума в зависимости от
возраста
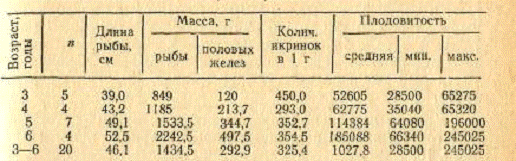
Кутум имеет категорию и статус 2 сокращающийся в
численности подвид вырезуба. В Северном Каспии кутум всегда был редок, наиболее
часто встречался в водах Азербайджана, где его уловы в 1921-1937 гг. достигали
360-2520 т или 200 1400 тыс. штук. В Тереке в прошлом был многочислен, где его
добывали до 80 т, в Волге заходило небольшое количество (в 1937-1940 гг. в Каме
было поймано всего 10 экз.), единичные экземпляры встречались в устье Урала.
Причины снижения численности зарегулирование стока рек, сокращение естественного
воспроизводства. В 1970-1973 гг в Азербайджане уловы не превышали 80 т, с конца
1070-х по всему Каспию кутума вылавливалось очень мало. Современных данных о
состоянии запасов кутума нет. Необходимо оценить численность кутума в
прибрежных водах Каспия на участке Самур-Терек. Меры охраны расширить работы по
искусственному разведению кутума до производственных масштабов (Красная книга
Российской Федерации, 2001).
.3 Размножение. Половые циклики и гаметогенез
кутума
Кутум - проходная рыба, большую часть жизни
проводит в море и только для икрометания идет в реки и речки, впадающие в
заливы Горган и Пехлевийский, в pp. Каладагна, Ленкорань, Кура, Кейранчай,
Кусарчай, Гильгильчай, Самур, Терек, единично в - pp. Сефидруд, Атрек, Гюргян,
Волга, Урал (Абдурахманов, 1982).
Кутум нерестится в медленно текущей воде
предустьевых участков рек на самом различном субстрате: на свежезалитой
растительности, в зарослях камыша, на подмытых корневищах прибрежной
растительности, а также на каменистом грунте. Характер развития икры и
поведение личинок кутума отличаются от таковых у вырезуба. Личинки кутума
подвешиваются с помощью специальных органов к растениям в тихих, заросших
участках реки, куда их заносит течение.
Половозрелости достигает в возрасте 4 лет, в
небольшом количестве, главным образом самцы, нерестится в возрасте 3 лёт.
Основную массу ходового кутума составляют рыбы в возрасте 5-6 и даже 7 лет
(Державин, 1951; Абдурахманов, 1962).
По сведениям А.Н. Державина (1934) и Н.И. Кожина
(1957), кутум, будучи в основном рыбой с полойным нерестом, имеет 2
биологические разновидности. Основная масса рыб идет для нереста в заливы
Аграханский Кирова и Пехлевийский, а также в приустьевые, заросшие камышом и
чаканом разливы их притоков (дельта р. Терека, pp. Самур, Кумбашинка и Сиазань)
другие популяции поднимаются в горные реки Талышского-Гилянского берега
(Ленкорань, Сефидруд) и мечут икру на галечном грунте как проходные рыбы
лососевого типа.
В дельте Терека кутум представлен половозрелыми
особями. В настоящее время нерест в основном происходит в сбросном канале, а
рыба зашедшая в залив, мечет икру преимущественно в его мелководных участках,
проточных и покрытых зарослями тростника и других растений. Часть
производителей через сбросной канал заходит в рыбоходный канал Новокосинского
нерестового хозяйства и нерестится в его прудах.
С конца января кутум начинает входить в
Аграханский залив главным образом через сбросной канал, частично-через
рыбоходный. Массовый ход в Аграханский залив наблюдается обычно в марте, к
концу апреля и началу мая он обычно завершается (Кулиев, Агаярова, 1964). Сроки
хода кутума зависят от условий года. Так, в начале 60-х годов отмечались очень
ранние сроки массового хода кутума: в 1960 г. нерестовый ход кутума начался в
первых числах февраля, в 1962 г. - в середине февраля, в 1964 г. - в начале
марта. Самый поздний массовый ход был зарегистрирован в 2004 году - в начале
апреля.
Особенности развития и роста кутума в раннем
онтогенезе исследованы Ш.М. Багировой (1967). По ее данным, в момент вылупления
из оболочки длина тела личинок колеблется от 14 до 17,5 мм, на 36-е сутки после
вылупления длина тела мальков составляет 19,0-33,0 мм, тело полностью
покрывается чешуей.
Кутум - быстрорастущая рыба. Молодь начинает
скатываться с конца мая и в начале июня, продолжается скат до конца июля.
Длина молоди, пойманной в третьей декаде июля у
полуострова Аргахан, колебалась от 15 до 60 мм, масса - от 1,4 до 1,7 г. С
конца июля количество молоди резко сокращается, в августе встречается очень
редко. Зависимость массы молоди от длины:
Длина, см 2,0 - 3,0 - 4,0 - 5,0 - 6,0 М
Масса, г 0,7 0,8 1,6 2,4 1,38
Рост молоди кутума в заливах Аграханском и
Кирова, по М.Ш. Багировой и 3.П. Бухариной (1967), характеризуется следующими
данными:
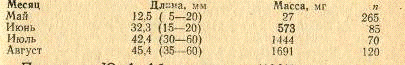
Но данным Ю.А. Абдурахманова (1982), молодь к
концу первого года достигает длины 6,8 см, массы 5,4 г. К концу третьего года
кутум вырастает в среднем до 32-39,6 см при массе 600-700 г, в возрасте 4 лет -
37-43 см и 800 г, 5лет - 41-49 см и 1200-1400 г, 6 -- 44-47 см и 1500- 1600 г,
7 -- 49-52 см и 2000 - 2200 г, 8 - 53-56 см и 2200-2700 г, 9-10-летние особи
достигают длины 59-62см.
Длина тела промыслового кутума, добываемого в
Каспии, колеблется от 34 до 55 см, в среднем 44,4 см, масса --от 685 до 2920 г,
в среднем 1570 г. Длина тела самок в среднем составляет 46,5 см, самцов - 42,3
см, масса - соответственно 1810 и 1825 г. Отдельные экземпляры достигают в
длину 67 см, массы 3910 г. Среди самок преобладают рыбы длиной 40-50 см
(70-80%), самцов - 35-45 см (70- 75%). В начале захода производителей в залив
рыбы имеют меньшие размеры, чем в период массового хода, к концу его масса рыб
уменьшается (Абдурахманов, Кулиев, Агаярова, 1968).
Таблица 8. Длина (числитель, см) и масса
(знаменатель, г) кутума из залива в различные годы
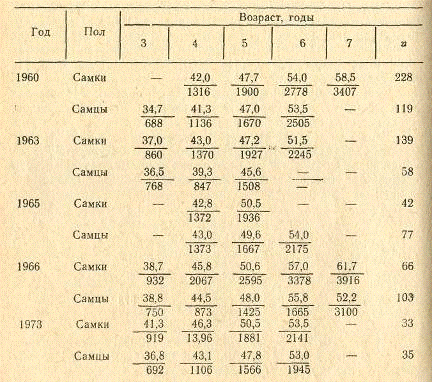
Данные непосредственного измерения кутума за
отдельные годы в заливе в преднерестовый период приведены в табл. 8. В уловах
встречаются рыбы в возрасте от 3 до 8 лет. Возрастной состав улова меняется по
годам (табл. 9). Основную массу улова за 1957-1958 гг. составляли рыбы в
возрасте 4-6 лет (Бердический, 1963) в 1960 г. (Абдурахманов, 1962) и 1965 г.
-- 4-5 лет. В 1963 г. нерестовая часть популяции в основном состояла из рыб
более молодых возрастных групп. Предельный возраст кутума в уловах - 11 лет.
Индивидуальный коэффициент упитанности (по Фультону) у кутума составляет
1,27-2,26, в среднем 1,74. Самки упитаннее самцов, что следует отнести за счет
развития их половых продуктов в весенний период. Это подтверждается сравнением
коэффициентов упитанности обоих полов по Кларк: у самок - 1,12-1,96, в среднем
1,54, у самцов - 1,20-2,10, в среднем 1,58. Коэффициент упитанности 10 самок,
пойманных в 1972 г., колебался (по Фультону) в пределах от 1,11 до 1,42, в среднем
1,22.
Таблица 9. Возрастной состав (в %) кутума в
промысловых уловах у полуострова Аграхан
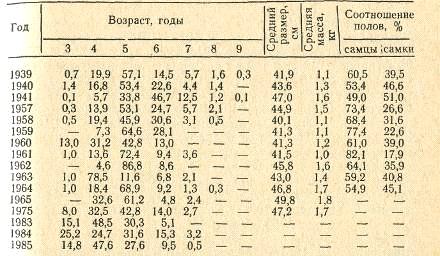
На ранних этапах развития кутум питается
коловратками, мелкими формами ветвистоусых рачков, диатомовыми водорслями,
циклопами, веслоногими рачками. Основными кормовыми объектами взрослых рыб
являются моллюски, меньше - амфиподы, креветки и др. (Абдурахманов, 1962). У
западного побережья Среднего Каспия кутум потребляет и крабов (Казанчеев,
1981).
Кутум - ценная промысловая рыба. Первоначально
значительная часть его уловов приходилось на Азербайджан. В 70-е годы заметно
увеличились уловы кутума в Среднем Каспии - вследствие проводимых в Дагестане
работ по воспроизводству стада этой рыбы, а в Южном Каспии, наоборот,
уменьшились в результате ухудшения условий воспроизводства его в районе залива
Кирова.
Микроскопическое (гистологическое) изучение
яичников и семенников позволило выявить ряд особенностей в развитии половых
клеток в течение годичного полового цикла. Гистологическая картина каждой
стадии зрелости у самок и самцов характеризуется наличием определенного
комплекса половых клеток, находящихся в разных фазах их роста и развития.
Переход половых желез от одной стадии к другой (от низшей к высшей)
определяется переходом половых клеток от одной фазы их развития к другой. Так,
для первой стадии зрелости яичников для всех видов рыб характерны образования
овоцитов ювенальной фазы развития и наличие первичных половых клеток -
овогоний.У самцов на этой стадии половые образования представлены первичными
половыми клетками - сперматогониями.
В яичниках второй стадии зрелости основная масса
овоцитов находится в фазе однослойного фолликула, отличающегося от овогоний
большим размером и характерной для премейо-тических фаз зрелости- структурой
ядра. Параллельно с процессом созревания овоцитов и переходом в последующие
фазы происходит заметное уменьшение количества первичных половых клеток -
овогоний. В семенниках второй стадии преобладают сперматоциты 1 порядка и
первичные сперматогонии.
Для третьей стадии характерно начало
вителлогенеза, т. е. большинство овоцитов находятся в фазе первоначального
накопления желтка (фаза вакуолизации); кроме того, имеется и комплекс овоцитов,
характерных для предыдущих II и I стадий У самцов на III стадии половые клетки
в семенниках представлены главным образом сперматоцитами второго порядка, но
имеются сперматоциты первого порядка и единичные сперматого-нии, характерные
для младших стадий зрелости.
При IV стадии зрелости яичников большая часть
овоцитов находится в фазе наполненного желтком, в остальной массе-овоциты всех
фаз развития, обнаруживаемые в предыдущих стадиях. Однако эта стадия имеет ряд
особенностей, свойственных как для единовременно, так и порционно нерестующих
рыб. Так, у единовременно нерестующих в яичниках на этой стадии имеются овоциты
в фазе наполненного желтком и комплекс овоцитов, присущих для II и I стадий
зрелости, но нет овоцитов, характерных для III стадии зрелости (в фазе
первоначального накопления желтка или вакуолизации),-тогда как у порционно
нерестующих рыб в яичниках имеются овоциты всех фаз развития, характерные для
предыдущих III, II и I стадий зрелости, т. е. овоциты во всех фазах
трофоплазматического и протоплазматического роста (рис. 1Д). У последних в
течение всего годичного полового цикла наблюдается асинхронность в росте и
развитии, которая стойко сохраняется в дальнейшем.
В семенниках в IV стадии зрелости семенные
ампулы заполнены сперматидами, которые при наступлении нерестовых температур за
короткий период переходят в зрелое состояние, т. е. в сперматозоиды.
Гистологическая картина пятой стадии по
некоторым внешним признакам сходна с IV, однако появляются некоторые признаки,
свойственные только для яичников пятой - предовуляционной стадии, а именно:
овоциты достигают дефинитивных размеров (зреяое состояние), в них появляются
«микропиле» с замыкающей клеткой (рис. 1Ж), являющейся местом проникновения
сперматозоидов для оплодотворения. В семенниках семенные ампулы приобретают
вытянутую или округлую формы, заполненные сперматозоидами, готовыми при
наступлении оптимальных условий к размножению и сразу же к выделению. У самцов
некоторых видов рыб (вобла, лещ, окунь, жерех, щука, рыбец и т. д.) сперма
выделяется при самом незначительном соцрикосновении с предметами в период
нерестовой миграции.
Гистологическая картина яичников и семенников в
посленерестовой VI стадии представляет интерес. У единовременно нерестующих рыб
после икрометания (овуляции зрелых овоцитов) в яичниках остаются лишь
единичные, невыметанные по разным причинам, икринки, многочисленные
фолликулярные оболочки, которые подвержены процессу резорбции, а также имеются
и овоциты протоплазматического роста. Вителлогенез начинается у них только по
завершении резорбционных процессов или даже при .наличии конечных фаз резорбции
в виде включений пигментных клеток.
У самцов одновременно с резорбцией остаточных
спермиев на стенках семенных ампул (канальцев) появляются первичные половые
клетки - сперматогонии (крупного размера), которые обычно расположены в них в
один ряд, в виде цепочки.
У самок кутума, (с единовременным нерестом)
яичники к осени переходят в IV стадию (интенсивный вителлогенез у них
наблюдаются с сентября по ноябрь) и зимуют они в этом состоянии половых желез.
Анализ сезонного хода полового цикла этих видов рыб показал возможность
получения от них зрелых половых продуктов значительно раньше, при создании
оптимальных условий для икрометания, чем до их естественного нереста.. Эту
возможность - вызывать в более ранние сроки овуляцию и оплодотворение -
непременно следует использовать как при искусственном рыборазведении, так и для
разведения этих видов рыб в естественных водоемах с коротким вегетационным
периодом и с неблагоприятными гидрологическими и гидрохимическими условиями в
них, особенно в преднерестовый и нерестовый период. У самцов кутума
сперматогенез завершается только весной перед нерестом, а самцы их зимуют с
половыми железами в переходной II-III или в III стадии зрелости. Время
прохождения и длительность отдельных стадий зрелости гонад самок и самцов в течение
годичного полового цикла у разных видов рыб, независимо от их близости в
систематическом и биологическом отношениях, заметно различаются. По данной
схеме можно легко определить сроки начала и продолжительность каждой стадии
зрелости как яичников, так и семенников указанных в ней различных видов рыб,
поэтому она имеет не только теоретическое значение, но может быть использована
и в практических целях, в частности, при искусственном рыборазведения.
По отличительным свойствам ово- и
сперматогенеза, срокам прохождения половых циклов и длительности отдельных
стадий зрелости все изученные виды рыб относят к двум различным группам. Так,
самки воблы, леща, щуки, кутума, жереха, окуня и судака входят в группу с
ранневесенним кратким нерестом. Длительность отдельных стадий зрелости этих
видов рыб более или менее сходна.
Вторая стадия у этих видов рыб длится около трех
месяцев, III стадия - 1,5-2 месяца, 1Устадия - 5-5,5 месяцев, послене-рестовая
VI стадия - 1,5-2 месяца. Зимуют самки с яичниками в IV стадии зрелости (с
завершенным вителлогенезом). Сроки начала и завершения каждой стадии у
отдельных видов неодинаковы. Так, переход яичников во II стадию у щуки, окуня,
кутума, жереха происходит в конце мая, а у воблы и леща - в конце июня, III
стадия у первых наступает в конце августа, а у вторых - в конце сентября. Такая
же картина наблюдается и при прохождении других стадий.
Опыты по искусственному разведению кутума начаты
в 1924 г. на р. Кумбашинка в Азербайджане и на Самурской рыбоводной станции в
Дагестане (Борзенко, 1926). В более широком масштабе они были повторены после
50-х годов работниками Дагрыбвода, Севкаспрыбвода и ВНИРО на Самурском
рыбоводном заводе (Берлянд, 1957) и сотрудниками Института зоологии АН
Азербайджанской ССР в Малом заливе (Аббасов, Агаларов, 1962; Аббасов, 1972).
Экологические условия жизни кутума на всех
этапах развития хорошо изучены, разработана биотехника его разведения, что
позволяет осуществлять интенсивное заводское воспроизводство этой рыбы. В 1980
г. впервые такие работы были осуществлены в Азербайджане в специально
построенном на берегу Малого залива инкубатории мощностью 50 млн. личинок в
год. Такое количество личинок получают ежегодно и в месячном возрасте выпускают
в море. Однако выпуск еще не окрепших личинок непосредственно в морскую воду
вряд ли можно признать эффективным. Целесообразно было бы сначала подращивать
их в садках или специально отгороженных участках и лишь после этого выпускать в
море. Именно это и используется дагестанскими рыбоводческими хозяйствами,
которые учли несколько неудачный опыт азербайджанских хозяйств.
Искусственное разведение кутума должно стать
основным путем поддержания численности этой ценной промысловой рыбы в бассейне
Каспийского моря.
Вырезуб и кутум - ценные рыбы. Их мясо обладает
высокими вкусовыми качествами. В связи с этим большое значение приобретает
искусственное разведение вырезуба и кутума на рыбоводных заводах. Кутум
особенно высоко ценится в Иране, где он употребляется в вяленом виде и идет на
приготовление различных национальных блюд.
Глава IV. Разведение кутума
Ещё в конце 50-х годов группа
научных работников ВНИРО под руководством Т. Берлянда разработала метод
искусственного разведения карповых, в первую очередь вырезуба и кутума. Во
время хода рыбы вылавливают производителей, отсаживают в садки раздельно самцов
и самок и выдерживают в них до созревания половых продуктов. От производителей
со зрелыми половыми продуктами икру и сперму отцеживают обычным способом, икру
искусственно осеменяют и отмывают от клейкости в чистой, часто сменяемой воде в
течение 2 ч.
Производителей кутума
заготавливают с середины марта до конца апреля с помощью стационарных ловушек -
забоек.
Пойманных самок и самцов
выдерживают в деревянных или сетчатых садках размером 3X2X1,5 м. Срок
выдерживания обычно не превышает 15 ч. Потомство получают при естественном
нересте или заводским способом. Аналогично можно получать потомство и
родственной формы кутума - вырезуба.
При использовании естественного
нереста производителей помещают в маточные пруды, соединенные с нерестовыми
канавками, куда рыбу пускают при наступлении нерестовых температур. В канавках
происходит икрометание. Рыбы откладывают икру на гальку на дне канавок. После
нереста производителей переводят обратно в маточные пруды. Личинок после
выклева помещают в личиночный садок, а затем в выростной водоем.
При заводском методе получения
потомства икру, полученную методом отцеживания, осеменяют сухим способом. После
отмывки икры от слизи, ила и клейкости её помещают в инкубационные аппараты
Ющенко. Но преимущественно размножение кутума происходит в условиях
внезаводского метода инкубации. Внезаводской метод инкубации икры осуществляют
двумя способами: инкубацией икры на субстрате и в аппаратах.
Клейкую икру весенненерестующих
рыб инкубируют на субстрате. Субстратом могут служить ветки ели, можжевельника
и лиственных деревьев, которые сначала замачивают в воде, а затем связывают в
небольшие венички и помещают в корыто с водой. Слой воды над помещенными в
корыто веничками должен быть 15-20 см. Оплодотворенную, но не отмытую от
клейкости икру рассеивают небольшими порциями на венички. Икра, попадая в воду,
выделяет клейкое вещество и прочно прикрепляется к субстрату.
Затем венички привязывают
комлем вверх к веревке, концы которой закрепляют за колья, вбитые на участке
реки глубиной 1 - 1,5 л, и опускают их в воду. Расстояние от дна должно
быть-15-20 см.
Такой способ инкубации икры
прост, но лишает возможности, организовать уход за икрой. Это приводит к тому,
что икринки заиляются, поражаются сапролегнией (зарастают гифами низших грибов
- Saprolegnia и Achlya) и многие из них погибают. Кроме того, этот способ не
позволяет точно учесть количество мертвых икринок, съеденных икринок и
количество выклюнувшихся эмбрионов.
Способ инкубации икры в
аппаратах, установленных в водоеме, является более эффективным, чем способ
инкубации икры на. субстрате. Однако и при нем процесс инкубации полностью
зависит от влияния гидрологических условий водоема.
Отмытую икру помещают в
аппараты Сес-Грина. Продолжительность инкубации икры в аппаратах, как и в
естественных условиях, определяется температурой воды. При температуре воды 8°
С инкубация продолжается около 18 суток. Выклюнувшихся личинок выдерживают в
тех же аппаратах Сес-Грина 5-8 суток и затем либо выпускают в реку, либо
высаживают в выростные водоемы - небольшие непроточные неглубокие пруды.
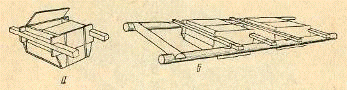
Рис. 3. Аппарат Сес-Грина:
а - общий вид; б - аппараты,
установленные в деревянную раму.
Аппарат Сес-Грина - деревянный
прямоугольный, ящик (длина 60 см, ширина 40 см, высота 25 см). Дно ящика
обтянуто металлической сеткой, покрытой асфальтовым лаком для предохранения от
ржавления (окисления). Размер ячеи сетки, должен быть меньше диаметра
инкубируемой икры. К торцовым; стенкам аппарата приделаны длинные деревянные
ручки, позволяющие поставить его в одно из гнезд деревянной рамы-плота,
установленной при помощи якорей на участке водоема с умеренной скоростью
течения (0,5-1,0 м/сек) и защищающей от волны, плывущих бревен и других
предметов. Вода свободно проникает через сетчатое дно аппарата и омывает икру.
Для удобства ухода за икрой внутри рамы-плота сделаны мостики, по которым можно
подойти к каждому аппарату.
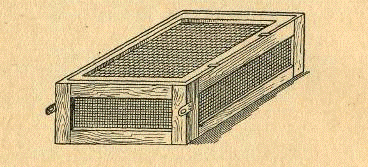
Рис. 4. Аппарат Чаликова.
Аппарат Чаликова представляет
собой ящик размером 70 X 34 X 15,5 см. Стенки аппарата состоят из деревянных
рамок, обтянутых металлической сеткой, покрытой асфальтовым лаком. Сверху ящик
закрывается сетчатой крышкой. В торцовые стенки ящика ввинчены металлические
кольца или прикреплены ушки, при помощи которых аппараты соединяют один с
другим.
Так как в аппарате Чаликова все
стенки обтянуты сеткой, в нем происходит лучший водообмен, а следовательно,
икра лучше омывается водой, чем в аппарате Сес-Грина. Второе преимущество
аппарата Чаликова то, что его можно погружать на любую глубину. Это позволяет
избежать случайного отравления икры плавающими на поверхности воды ядовитыми
веществами, например нефтью.
Нормы загрузки икры в аппараты
Сес-Грина и Чаликова зависят от размеров аппаратов, условий водообмена,
температуры воды и содержания в ней кислорода. Водообмен зависит от скорости
течения. Поэтому нормы загрузки аппаратов икрой зависят от района и периода
инкубации. Чем ниже температура и сильнее течение воды, тем выше нормы загрузки
аппаратов икрой, и наоборот.
Примерные нормы загрузки икры в
аппараты Сес-Грина и Чаликова следующие: севрюги-15-35 тыс., леща-125- 200
тыс., судака - 200-250 тыс. шт.
Личинки кутума в возрасте 5-б
суток начинают питаться зоопланктоном (низшие ракообразные, коловратки). При
достижении размеров 20 мм они переходят на питание придонными и донными
организмами, преимущественно личинками хирономид.
При искусственном разведении
кутума следует пользоваться следующими расчетными показателями:
. Рабочая плодовитость 60-65
тыс. икринок.
. Норма загрузки в аппараты
Сес-Грина:
-70 тыс. икринок при диаметре
ячеи сетки 0,9-1,0 мм и скорости течения воды в аппарате 0,4-0,7 м/сек;
-35 тыс. икринок при диаметре
ячеи 0,5 мм, скорости течения 0,4 м/сек.
. Отход икры за период
инкубации 30%.
. Отход личинок при
выдерживании (4-5 суток) 15%.
. Норма посадки личинок в
выростные водоемы: при монокультуре-100-120 тыс. шт/га; при совместном
выращивании с лещом - 20-30 тыс. шт/га.
. Выживание за период
выращивания личинок в выростных водоемах 70%.
. Средняя масса молоди кутума
(в возрасте 3 месяцев) 1,2- 1,5 г.
В последние годы на НВВ
проведены работы по повышению эффективности воспроизводства ценных промысловых
рыб: реконструированы Аракумские водоемы, мелиоративные работы в Каракольском
водоеме позволили решить проблему водообеспечения и проводить весной
необходимый сброс воды по рыбоходу для привлечения производителей. В результате
скат молоди в последние годы заметно увеличивается и достиг в 2002 году 60 млн
шт. (таблица 10) Важно отметить, что в последние годы наблюдается повышение
роли НВВ дельты р. Терека в естественном воспроизводстве таких видов, как кутум
и рыбец. По существу кутум, рыбец и шемая были на грани исчезновения как
биологические виды, но именно благодаря НВВ их численность возросла, и в
настоящее время кутум имеет промысловое значение. Это одна из наиболее ценных
промысловых рыб Каспийского моря. В прошлом его запасы на дагестанском
побережье поддерживались за счет естественного размножения в пойменных озерах
дельты Терека и родниковых ручьях дельты Самура и нагула в Каспийском море,
промысловые уловы достигали 600-800 т в год.
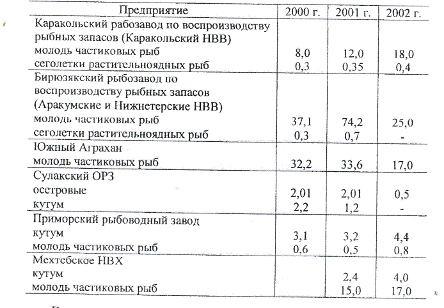
Таблица 10. Воспроизводство
молоди ценных видов рыб в Дагестане в 2000-2002 гг., млн. шт.
Основным мероприятием, которое
должно было обеспечить увеличение запасов кутума на Каспии, являлось создание
благоприятных условий воспроизводства. Решить эту задачу можно было
строительством нерестово-выростных водоемов. Специально для воспроизводства
кутума был построен Самурский НВВ, представляющий собой обвалованный участок
дельты Самура площадью 100 га. После создания НВВ запасы кутума стали
восстанавливаться промысловые уловы. По средним многолетним данным, в Самурский
НВВ на нерест заходило от 100 до 200 тыс. производителей и скатывалось в море
10-15 млн шт. молоди. Из водоемов дельты Терека в море скатывалось 3,5-4,0 млн
шт. молоди кутума. Имеющиеся запасы позволили организовать его промышленную
добычу. В 1983 г. было выловлено 41 т, а в 1984 г. - 80 т кутума.
Однако с поднятием уровня моря
была размыта оградительная дамба Самурского НВВ и площадь водоема уменьшилась
до 16 га. Скат молоди снизился до 3,0-3,5 млн шт. В водоемах дельты Терека
из-за трудностей с водообеспечением воспроизводство кутума прекратилось и с
1998 г. он был занесен в Красную книгу.
В настоящее время возникла
необходимость организации заводского воспроизводства кутума. В 1998 г. в
Ассоциации «Дагрыба» была разработана Программа искусственного воспроизводства
кутума в Дагестане на период до 2005 г., где предполагается использовать
производственные мощности Бирю-зякского рыбопитомника. В 2000 г. в питомнике
выращено более 0,5 млн шт., а к 2005 г. ожидается 4,0 млн. шт. Реализация
программы позволит в ближайшие годы восстановить и поддержать численность вида,
обеспечивающую объем его промысла до 200 т.
Одновременно следует
регулировать состояние водоёмов, где производится разведение кутума.
Рыбопродуктивность рыбохозяйственных водоемов (озер, водохранилищ, прудов и
рек), как и плодородие сельскохозяйственных угодий, не остается постоянной. Она
колеблется во времени и зависит от состояния водоемов: степени заиленности и
характера иловых отложений, газового и температурного режима, плотности и
характера размещения высшей мягкой и жесткой водной растительности. Сказывается
на рыбопродуктивности водоема и его водный режим: постоянен ли уровень воды в
водоеме или характер его колебания то увеличивается, то резко падает. Во многом
она зависит и от системы эксплуатации водоема, формы организации рыбного
хозяйства. Чтобы обеспечить высокую и устойчивую рыбопродуктивность водоемов,
рыбоводы применяют искусственные биотехнические приемы для улучшения
физико-химических и гидробиологических условий водоемов (удаление коряг,
излишков ила и водной растительности).
При недостаточности иловых
отложений и отсутствии необходимых солей в воде вносят органические и
минеральные удобрения и регулируют сток, при недостатке кислорода в воде -
аэрируют.
Система мероприятий,
направленных на улучшение биохимических и эксплуатационных качеств водоемов в
рыбном хозяйстве, получила название рыбохозяйственной мелиорации.
По характеру и
продолжительности действия на водоем мелиоративные мероприятия подразделяются
на коренные и текущие. Коренные мелиоративные мероприятия требуют больших
затрат и оказывают свое действие в течение ряда лет. Текущая мелиорация
оказывает свое действие в течение одного года, точнее- вегетационного периода.
Многообразие естественных факторов, обусловливающих рыбопродуктивность
водоемов, порождает разнообразие и видов мелиоративных работ.
В отношении водоёмов дельты
Терека полезно было бы проводить мелиоративные работы по предупреждению
заиления и зарастания водоёмов. Этот вид мероприятий относится к комплексу,
обеспечивающему улучшение условия питания и жизни рыб в водоеме. Заиление
водоема происходит в результате действий различных факторов: эрозии, оседания
остатков водных растений и животных, отмерших в самом водоеме,
жизнедеятельности аэробных бактерий, населяющих верхний слой ила, разлагающих
остатки растений и животных и придающих в результате своей жизнедеятельности
характерную консистенцию и состав илу.
Быстрота заиления водоема
зависит от топографических и физических особенностей площади водосбора и
состава его почв. Отсюда следует, что для замедления процесса заиления водоемов
надо прежде всего предотвратить смыв почв с площади водосбора. Этого можно
достигнуть путем устройства защитных лесных и луговых полос вокруг водоема и
вдоль берегов рек. Слой иловых отложений свыше 20-25 см снижает биологическую
продуктивность водоема и затрудняет вылов рыбы. Заиленность водоема
способствует развитию высшей мягкой и жесткой растительности, зарастанию и
заболачиванию водоемов.
Из мягкой водной растительности
в наших водоемах обычны тысячелистник, роголистник, разные виды рдестов; из
жесткой надводной растительности - тростник, камыш, рогоз, аир. Жесткая
подводная растительность вначале появляется в мелководной части, окружает
водоем, постепенно продвигается к его центру и покрывает всю площадь водоема.
Огромно значение водной растительности
и в жизни рыб. Небольшие заросли водных растений являются местом нереста и
нагула молоди промысловых рыб, а массовое развитие становится неблагоприятным
фактором и требует вмешательства рыбоводов в целях его ослабления.
Обычно жесткую водную растительность
выкашивают обыкновенными косами и разного типа механизированными
камышекосилками. Мягкую водную растительность (в том числе и ряску) удаляют из
водоема специально сконструированными небольшими бреднями или колючей
проволокой. Для удаления мягкой растительности применяется водяная борона
(рис.) -деревянный треугольник с зубьями из гвоздей с прикрепленными по углам
крюками. Чтобы борона не всплывала и плотно держалась на дне, ее загружают
грузом. Веревкой, прикрепленной к одному из углов, борону протаскивают по пруду
и таким путем извлекают мягкую растительность. Вместе с этим рыхлится верхний
слой грунта, что усиливает в нем аэробные процессы.
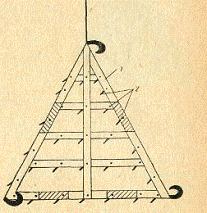
Рис. 5. Водяная борона для
удаления мягкой водной растительности и взрыхления поверхности ила:
- деревянная рама; 2 - зубья.
Наряду с механическими
способами удаления растительности используют и химические средства (гербициды),
но только безвредные как для рыб, так и для фитопланктона и фауны
беспозвоночных водоема. В РФ и США в этих целях применяют медный купорос
(CuSCy5H2O) и мышьяковистый натрий. В последнее время для борьбы с зарослями
тростника рекомендуется бутиловый эфир 2,4Д. Медный купорос применяется главным
образом для уничтожения водорослей. Для этого раствор его в концентрации 0,33
мг/л вносится в водоем путем разбрызгивания. Следует помнить, что и медный
купорос при наличии в воде карбоната кальция становится токсичным для рыб.
Мышьяковистый натрий в растворе
1,7-4 мг/л используют главным образом для борьбы с цветковыми растениями. В
такой концентрации для рыб он нетоксичен. Слабо действует он также на водную
лилию и надводную растительность.
Бутиловый эфир применяется в
форме эмульсии в концентрации 1,25 мг/л.
Опрыскивать тростник
рекомендуется на ранней стадии развития, до выбрасывания соцветий.
Бутиловый эфир в эмульсии
концентрацией 0,005 г/л для рыб нетоксичен.
Работать с ядохимикатами надо
осторожно: в спецодежде (или по крайней мере в фартуке), специальных перчатках
и обязательно в защитных очках.
Для борьбы с заилением и
зарастанием водоемов можно пользоваться агрорыбоводным методом, предложенным А.
Т. Болотовым ещё в 1785 г., а впоследствии незаслуженно забытым. Этот метод
заключается в том, что водоем периодически (в межрыбоводный период) спускают, дно
просушивают, вспахивают и используют под посев трав и других
сельскохозяйственных культур. В настоящее время такое своеобразное чередование
выращивания рыбы с засевом ложа водоема сельскохозяйственными культурами в
межрыбоводный период начинает постепенно вводиться, например, на осетровых
заводах, нерестово-выростных хозяйствах. Это повышает рыбопродуктивность
водоемов и значительно снижает, а иногда и полностью ликвидирует их зарастание
водной растительностью.
Комплекс мероприятий по
улучшению условий эксплуатации водоемов в основном сводится к ликвидации
причин, мешающих вылову рыбы, улучшению условий размножения рыбы и сохранению
молоди.
При определении масштабов работ
по ликвидации причин, мешающих вылову рыбы, составляется карта-схема водоема,
согласно которой намечается план мероприятий. Размеры и порядок этих работ
зависят от состава ихтиофауны и ее распределения в водоеме в разные сезоны.
Если рыба держится на засоренных, закоряженных участках, разбросанных по всему
водоему, то необходимо расчистить весь водоём. Если основная масса промысловой
рыбы приурочена к каким-то определенным местам, ограничиваются мелиорацией
только этих участков.
К мероприятиям по улучшению
условий размножения рыбы и сохранению молоди относятся:
· сохранение и
улучшение естественных нерестилищ;
· устройство
искусственных нерестилищ;
· спасение молоди из
отшнуровывающихся полоев.
В связи с гидростроительством
наибольшее значение имеет устройство искусственных нерестилищ и спасение молоди
ценных промысловых рыб.
Донные устраиваются на
мелководьях в озерах и водохранилищах. Для этого выбирают места, освещаемые
солнечными лучами и защищенные от господствующих ветров. На дне намеченных
участков размещаются и укрепляются ветки ели, можжевельника, а иногда и
лиственных пород.
Такое искусственное нерестилище
дает рыбоводу возможность контролировать нерест и учитывать выход молоди.
Плавучие нерестилища,
конструкции сотрудников ВНИИПРХа (Б. Себенцов, П. Михеев и Е. Мейснер),
применяются в водохранилищах и некоторых озерах. Основу такого нерестилища
составляет рама из деревянных жердей, ширина которой 1 м, а длина зависит от
гидрологических условий в месте установки. К раме через 30-40 см привязывают
поводки из мочала или веревки длиной 3-6 м. К поводкам через 30-40 см,
привязываются венички из веток елки, можжевельника, промытых корневищ тростника
или камыша с таким расчетом, чтобы верхний веничек располагался на 0,5 м ниже
поверхности воды, а нижний отстоял на 0,25 м от дна.
Такая система плавучих
искусственных нерестилищ устанавливается вблизи подходов к естественным
площадям. В целях охраны плавучего нерестилища и особенно защиты икры
рекомендуется каждую нерестовую конструкцию после окончания икрометания
ограждать мелкоячейной делью или устраивать специальные плавучие садки, куда
следует помещать венички с икрой для инкубации.
Сохранение и улучшение
естественных нерестилищ осуществляются путем расчистки и углубления протоков и
ериков, связывающих полой с рекой, углубления основных путей, по которым рыба
переходит из авандельты, в дельту. В период паводка регулируют водный режим
полоев путем устройства обвалований с низовой стороны отдельных участков дельты
и на протоках (для образования подпора воды) и системы шлюзов для регулирования
стока.
Заключение
Основные выводы по результатам исследования:
.Кутум заходит в Терек в середине марта
(отклонения по времени зависят от условий в каждом данном году), нерестится в
феврале-мае, икру выметывает на заросли камыша в разливах рек и в озерах. Как и
вырезуб, кутум созревает на 4-5-м году жизни.
.Кутум нерестится в медленно текущей воде
предустьевых участков рек на самом различном субстрате: на свежезалитой
растительности, в зарослях камыша, на подмытых корневищах прибрежной
растительности, а также на каменистом грунте. Характер развития икры и поведение
личинок кутума отличаются от таковых у вырезуба. Личинки кутума подвешиваются с
помощью специальных органов к растениям в тихих, заросших участках реки, куда
их заносит течение.
3.Кутум, будучи в основном
рыбой с полойным нерестом, имеет 2 биологические разновидности. В дельте Терека
кутум представлен половозрелыми особями. В настоящее время нерест в основном
происходит в сбросном канале, а рыба зашедшая в залив, мечет икру
преимущественно в его мелководных участках, проточных и покрытых зарослями тростника
и других растений.
Основные предложения по результатам
исследования:
1.Выпуск еще не окрепших
личинок непосредственно в морскую воду вряд ли можно признать эффективным.
Целесообразно было бы сначала подращивать их в садках или специально отгороженных
участках и лишь после этого выпускать в море. Искусственное разведение кутума
должно стать основным путем поддержания численности этой ценной промысловой
рыбы в бассейне Каспийского моря.
.Чтобы обеспечить хорошее
развитие рыбы, корма должны содержать в определенном соотношении основные
питательные вещества - белки (протеины), жиры и углеводы, а также минеральные
соли и витамины. Неполноценность отдельных кормов устраняется путем составления
рационов из комплекса различных кормовых компонентов. В такой комплекс должны
входить в необходимом количестве все питательные вещества, в которых нуждается
организм рыбы.
.Следует регулировать состояние
водоёмов, где производится разведение кутума. В отношении водоёмов дельты
Терека полезно было бы проводить мелиоративные работы по предупреждению
заиления и зарастания водоёмов. Этот вид мероприятий относится к комплексу,
обеспечивающему улучшение условия питания и жизни рыб в водоеме. Для борьбы с
заилением и зарастанием водоемов можно пользоваться агрорыбоводным методом,
предложенным А. Т. Болотовым ещё в 1785 г. Он заключается в том, что водоем
периодически (в межрыбоводный период) спускают, дно просушивают, вспахивают и
используют под посев трав и других сельскохозяйственных культур.
.К мероприятиям по улучшению условий
размножения рыбы и сохранению молоди относятся: сохранение и улучшение
естественных нерестилищ; устройство искусственных нерестилищ; спасение молоди
из отшнуровывающихся полоев. Сохранение и улучшение естественных нерестилищ
осуществляются путем расчистки и углубления протоков и ериков, связывающих
полой с рекой, углубления основных путей, по которым рыба переходит из
авандельты, в дельту. В период паводка регулируют водный режим полоев путем
устройства обвалований с низовой стороны отдельных участков дельты и на
протоках (для образования подпора воды) и системы шлюзов для регулирования
стока.
Список использованной литературы
1. Водный
кодекс Российской Федерации от 16 ноября 1995 г. N 167-ФЗ (с изм. и доп. от 30
декабря 2001 г., 24 декабря 2002 г., 30 июня, 23 декабря 2003 г.). Федеральный
закон от 31 июля 1998 г. N 155-ФЗ "О внутренних морских водах,
территориальном море и прилежащей зоне Российской Федерации" (с изм. и
доп. от 22 апреля, 30 июня, 11 ноября 2003 г.).
2. Федеральный
закон от 8 декабря 1995 г. N 193-ФЗ "О сельскохозяйственной
кооперации" (с изм. и доп. от 7 марта 1997 г., 18 февраля 2003 г., 21
марта 2002 г., 10 января, 11 июня 2003 г.).
. Федеральный
закон от 30 ноября 1995 г. N 187-ФЗ "О континентальном шельфе Российской
Федерации" (с изм. и доп. от 10 февраля 2003 г., 8 августа 2001 г., 22
апреля, 30 июня, 11 ноября 2003 г.)
. Федеральный
закон от 24 апреля 1995 г. N 52-ФЗ "О животном мире" (с изм. и доп.
от 11 ноября 2003 г.).
. Указ
Президента РФ от 29 августа 1997 г. N 950 "О мерах по обеспечению охраны
морских биологических ресурсов и государственного контроля в этой сфере"
. Постановление
Правительства РФ от 7 апреля 2004 г. N 182 "Вопросы Федерального агентства
по рыболовству".
. Постановление
Правительства РФ от 4 июля 2002 г. N 496 "Об утверждении Положения о
лицензировании осуществляемой в море деятельности по приемке и транспортировке
уловов водных биологических ресурсов, включая рыб, а также других водных
животных и растений"
. Постановление
Правительства РФ от 9 декабря 1998 г. N 1457 "О выплате в 1998 году
выходного пособия и компенсаций работникам органов рыбоохраны и воспроизводства
рыбных запасов Министерства сельского хозяйства и продовольствия Российской
Федерации"
10. Абдурахманов
Ю.Ф. Рыбы залива Кирова Баку: 1988 - 328 с.
11. Аннотированный
каталог круглоротых и рыб континентальных вод России // Под ред.
Ю.С.Решетникова. М.: Наука, 1998. 218 с.
. Атлас
пресноводных рыб России: В 2 т. Т.1. // Под ред. Ю.С. Решетникова. М.: Наука,
2004 - 379 с.
. Берг
Л.С. Рыбы пресных вод СССР и сопредельных стран. М.; Л.: Изд-во АН СССР. 1949
Т. 2. С. 469-929.
14. Биологические
ресурсы мирового океана” П.А. Моисеев., Москва., ”Пищепромиздат”, 1989
. Богуцкая
Н.Г. 1986. К вопросу о систематическом отношении видов родов Abramis, Blicca,
Vimba (Cyprinidae) //Вопр. ихтиологии. Т. 26, вып. 4. С. 576-584.
16. Васильев
В.П. Эволюционная кариология рыб. М.: Наука, 1985. 300 с.
. Вилер
А. Определитель рыб морских и пресных вод М.: Инфра-М, 2003 - 457 с.
. Виноградов
В.К., Ерохина Л.В. Повышение эффективности товарного рыбоводства // Рыбоводство
и рыболовство - 2003.- № 3. - С. 6.
. Глущенко
В.Д. Рыбы в России должно быть больше // Рыбоводство и рыболовство. - 2003. - №
3. - С. 2-4.
. Долгий
В.Н. Ихтиофауна бассейнов Днестра и Прута. Кишинев: Штиинца. 2003 - 319 с.
. Иванов
А.П. Рыбоводство в естественных водоемах. - М.: Агропромиздат, 1988. - 368 с.
. Казанчеев
Е.Н. Рыбы Каспийского моря. М.: Пищепром, 1981 - 167 с.
. Кулиев
З.М. Рыбы Каспийского моря. Баку: 1988 - 224 с.
. Красная
книга Российской Федерации (животные). М.: АСТ, Астрель, 2005 - 860 с.
25. Охрана
окружающей среды в рыбном хозяйстве”/ под ред. Н.И. Осиповой М: Агропром, 2003
- 320 с.
. Павлов
Д.С., Савваитова К.А., Соколов Л.И., Алексеев С.С.. Редкие и исчезающие
животные: Рыбы. М.: Высшая школа, 1994 3- 34 с.
27. Промысловые
рыбы СССР. М.: Пищепромиздат, 1949. С. 407-409.
. Решетников
Ю.С., Богуцкая Н.Г., Васильева Е.Д. и др. Список рыбообразных и рыб пресных вод
России //Вопросы ихтиологии. 1997.Т. 37, вып. 6. С. 723-771.
29. Ройс
В.Ф. Введение в рыбохозяйственную науку М.:Пищепром, 1985 - 272 с.
30. Рыбоводство
/ авт.-сост. А.И. Исаев М.: Агропром, 1996 - 432 с.
31. Рыбы
Подмосковья. М.: Наука, 1988 - 141 с.
. Сорвачев
К.Ф. Основы биохимии питания рыб М.: Лёгкая и пищевая промышленность, 1988 -
247 с.
33. Справочник
по рыбоохране М.: Агропромиздат, 1986 - 288 с.
34. Троицкий
С.К., Цуникова Е.П.1988. Рыбы бассейнов Нижнего Дона и Кубани. Ростов н/Д:
Рост. кн. изд-во. 112 с.
35. Федорова
З.В. Настоящее и будущее аквакультуры // Рыбное хозяйство. Сер. Аквакультура:
Информационный пакет. - Вып. 4. - М.: ВНИЭОХ, 1998. - С. 1-7.
. Ферид-Пак
М. Морфология кутума Баку: 1992 - 271 с.
. Цепкин
Е.А. Изменения промысловой фауны рыб континентальных водоемов Восточной Европы
и Северной Азии в четвертичном периоде //Вопр. ихтиологии. 1995. Т. 35, вып. 1.
С. 3-17.
. Шутов
В.А. Ревизия рода Blicca и некоторые данные относительно филетических связей
между представителями рода Abramis (Pisces, Cyprinidae) // 1989 Зоологический
журнал, т. 48. С. 1105-1107.