Влияние скорости течения воды на рост колониальных гидроидов
Влияние скорости течения воды на рост колониальных
гидроидов (hydrozoa, thecaphora)
Н.Н. Марфенин
Имеются
разрозненные данные о влиянии подвижности воды на форму побегов у некоторых
гидроидов [1, 5, б, 7], но нет никаких сведений о скорости перемещений
окружающей воды, при которых колонии у разных видов лучше всего растут.
Многолетние
исследования различных видов гидроидов на Еремеевском пороге Ругозерской губы
Белого моря привели автора к предположению о наличии у них межвидовых различий
в гидродинамическом преферендуме, обусловленных особенностями морфологии
каждого вида. Экспериментальная проверка этого предположения — предмет данной
работы.
Исследование
проведено на Беломорской биостанции МГУ в июле-августе 1983 г. на четырех видах
гидроидов из подотряда Thecaphora: Obelia gemculata (L.), 0. longissima
(Pall.), 0. loveni (Allro.), из семейства Campanulariidae и Dynamena pumila
(L.), из семейства Sertulariidae. Все четыре вида близкородственны (три
принадлежат к одному роду и обитают на Еремеевском пороге в непосредственной
близости друг от друга в зоне нижней литорали — верхней сублиторали на
водорослях и камнях. Перемещение воды здесь происходит вследствие
приливно-отливных течений; поэтому оно характеризуется правильной
периодичностью, ламинарностью, регулярным изменением направления и постоянным
изменением скорости в пределах от 0 до 77 см/с.
Для
изучения влияния различных скоростей перемещения воды относительно объекта на
его рост использована разработанная Б.Я. Виленкиным с соавторами [3] простая
методика. Стекла с выросшими на них колониями или проросшими пла-пулами
укреплялись на перекладине, вращающейся с постоянной скоростью наподобие пропеллера
над круглым бассейном. В то время, как верхняя часть стекла, за которую стекло
прикреплено к "пропеллеру', была над водой, нижняя его часть с находящейся
на ней колонией постоянно оставалась в воде. Скорость перемещения стекла с
колонией относительно воды в бассейне зависела от расстояния крепления стекла
от центра (оси) "пропеллера". Реальная скорость обтекания объекта
водой представляет собой разность между линейной скоростью вращения стекла и
линейной скоростью вращения воды в бассейне на данном расстоянии от его центра,
которая определялась эмпирически.
В
четырех сериях опыта реальные скорости обтекания объекта водой были:
1—9
см/с; II -31 см/с; III -51 см/с; IV —75 см/с. что соответствует скоростям
течений в море.
Смену
воды в бассейне производили через день. Гидроидов кормили свежевылупившимися
науплиусами Artemia ежедневно, добавляя корм непосредственно в бассейн.
Концентрацию науплиусов периодически проверяли, отбирая пробы по 250 мл из
разных мест бассейна. В центре бассейна концентрация была иногда в 1,5 — 2 раза
выше, чем на его периферии, кроме того, она менялась повсюду в течение времени,
но в среднем была примерно 200—300 науплиусов на 1 л. Более высокая
концентрация корма в центре бассейна не мешала проведению исследования, так как
частота встречи с добычей понижается при уменьшении скорости обтекания колонии
водой. Исходя из этих соображений и расчетов частота встречи гидрантов с
жертвами оставалась на периферии при заданных скоростях вращения в 3—4 раза
выше, чем в центре бассейна.
Колонии
двух видов гидроидов (О. geniculata и O.longissima) получены из проросших на
стеклах фру стул (разновидность бесполого размножения [2]), колония O.loveni —
из планул, а колонии D.pumila - из изолированных взрослых побегов одинаковой
величины (8—10 междоузлии). В первом и последнем случае процесс выращивания
колоний относительно прост: их можно получать в любом количестве и по
собственному выбору размещать на субстрате. Выращивание же колоний на стеклах
из планул значительно сложнее из-за их подвижности. Удалось получить немного
пригодных для проведения эксперимента колоний, чем объясняется маленькая
выборка у O.loveni.
Регистрацию
морфологических изменений и роста колонии проводили методом картирования [4]
каждые вторые сутки.
Проведенный
эксперимент показал, что рост колонии одного вида различается в зависимости от
скорости обтекания их водой (табл. 1). На основании картирования колоний и
измерений протяженности ценосарка столонов и побегов высчитывался прирост
колоний за 8 суток. Для этого определяли во сколько раз общая протяженность
ценосарка колонии в конце опыта (на восьмые сутки) была больше, чем вначале (в
процентном выражении).
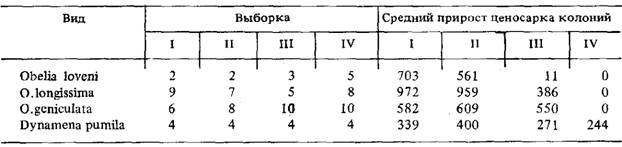
Величина
прироста колонии, как правило, убывает с возрастанием скорости потока воды
вокруг нее. У разных видов из числа изученных это выражалось в неодинаковой
степени (табл.1).
Так,
у O.loveni колонии лучше всего росли при скорости обтекания 9 см/с. Уже при
скорости обтекания, равной 31 см/с, происходило заметное снижение прироста, а
при еще больших скоростях (51 и 75 см/с) колонии фактически переставали расти.
У O.longissima и О, geniculata колонии переставали расти только при скорости
обтекания 75 см/с. Однако у O.longissima при скорости обтекания 51 см/с
происходит значительное снижение прироста колоний, а у О. geniculata этого не
наблюдалось. Наконец, ,у D.pumila ни при каких заданных скоростях обтекания
остановки роста колоний не было, хотя отмечено некоторое снижение прироста при
наибольшей и наименьшей скоростях обтекания.
Эти
данные позволяют установить гидродинамический преферендум исследованных четырех
видов гидроидов. Очевидно, что сравниваемые виды приспособлены к обитанию на
течениях с разными скоростями.
Механизм
воздействия скорости течения на рост колонии связан с пищевым поведением —
заякориванием, удержанием и проглатыванием добычи, проносимой мимо гидранта
потоком воды.
Основание
для такого предположения дают результаты эксперимента, в котором использованы
довольно большие колонии на стеклянных пластинках (длина ценосарка 20—80 мм)
трех видов гидроидов (за исключением О. genicuiata), находившихся большую часть
суток в аквариуме с профильтрованной через мелкоячеистый газ водой. Только на
время кормления (4 ч ежедневно) пластинки с гидроидами помещали в описанную
выше установку, где с помощью их вращения создавалось обтекание колоний водой с
заданными скоростями. Таким образом, большую часть времени колонии находились в
одинаковых условиях и только 4 ч в сутки условия между сериями опыта
различались по скорости обтекания колоний водой во время их кормления. В этом эксперименте
скорости обтекания были заданы несколько иные: 9, 27 и 55 см/с. Кроме того, в
каждой серии было лишь по одной колонии. Тем не менее результат получился
однозначный, такой же как и в основном эксперименте.
Таблица
1
Средний
прирост (%) ценосарка колоний 4 видов гидроидов за 8 сут при разных скоростях
обтекания их водой: 1-9 см/с, II - 31 см/с, III - 51 см/с, IV ~ 75 см/с. У
D.pumila прирост указан для гидроризы
Ориентация
колоний всех видов по отношению к направлению потока воды не влияла на их рост.
Полученные
в эксперименте данные позволяют яснее представить важную роль гидродинамики
водной среды в экологии гидроидов. Течение необходимо гидроидам для приноса
пищи. В то же время при сильном течении гидроиды голодают, так как осевые
побеги прижимаются сильным течением к субстрату, а боковые ветви — к стволу
побегов. В результате колония уже не перехватывает поток и не способна отловить
из него добычу.
Жесткость
конструкции побегов различна у разных видов гидроидов. Среди обсуждаемых видов
у O.loveni побеги самые слабые из-за малого диаметра трубки перисарка и тонких
его стенок. У O.longissima больше и диаметр трубки перисарка, и толщина стенок.
У O.geniculata междоузлия побегов более сложной, чем цилиндрическая, формы, за
счет чего возрастает жесткость побега. У D.pumila гидротеки прирастают
прилежащей стороной к стволу, что значительно увеличивает его жесткость. Таким
образом, морфология гидроидов и условия их эффективного питания объясняет
влияние скорости течения на рост колоний,
В
естественных условиях мельчайшие различия в морфологии колонии оказываются
определяющими адаптацию видов к различающимся по гидродинамике условиям
существования. Это объясняет сосуществование на близком расстоянии друг от
друга родственных видов гидроидов.
Полученные
в эксперименте результаты хорошо согласуются с наблюдениями Ведлера [8] по
распределению и форме колоний гидроидов. По его данным размеры побегов таких
гидроидов, как Chidoscyphus marginatus, Sertularella speciosa, тем меньше, чем
больше скорость течения в месте обитания колоний. Причем предельной скоростью
течения для них оказывается, так же как и в наших опытах, 70 см/с.
Автор
благодарен Б.Я. Виленкину за ценные советы, а А.В. Хоменко и В.Н. Курашеву за
помощь в проведении исследований.
Список литературы
1.
Бируля А. Ежегодник Зоол. музея Имп. Акад. наук, 1898, с. 203—212. 2.
Иванова-Квзас ОМ. Бесполое размножение животных. Л.: Изд-во ЛГУ, 1977, с.
1-240- 3. Виленкин Б.Я. и др. - Зоол. жури., 1981, т. 60, вып. 10, с.
1480-1485. 4. Марфенин Н.Н. В кн.: Теоретич. и практ. значение
кишечно-полостных. Л., 1980, с. 66—69. 5. Наумов Д.В. Гидроиды и тидромедузы
морских, солоноватоводных и пресноводных бассейнов СССР. М„ Л,: Иэд-во АН СССР,
1960, с. 1-585, табл. I-XXX. 6. Motz'Kossowska S. - Arch.
Zool. exp. gen., 1911, vol. 6, 5 Ser., p. 325-352. 7. Hughes R.G. In:
Developmental and cellular biology of coelenterates. Proc. IV Intern.
Coelenterate Conf.” Interlaken, 4-8 Sept., 1979, Amsterdam, 1980, p. 179-184.
8. Wedler E. - Helgoland. wiss.Meeresuntersuch., 1975, vol. 27, № 3, p.
324-363.
Для
подготовки данной работы были использованы материалы с сайта http://soil.msu.ru/