Эколого-физиологические особенности доминирующей водоросли литоральной зоны озера Байкал
Введение
Макроводоросли являются одним из основных
компонентов первичного звена экосистемы озера Байкал. Они оказывают влияние на
кислородный режим в литорали, служат пищей и убежищем для беспозвоночных
животных [37, 36, 53, 54].
В зависимости от глубины изменяются
флористический состав водорослевых сообществ, биомасса и фотосинтетическая
активность, образующих их видов, что во многом определяется динамикой
физических параметров среды в частности интенсивностью света, спектром
рассеянной радиации, температурой [111, 119,143, 114, 107, 110, 111, 103].
В первую половину прошлого века В.Ч.
Дорогостайским [17] и А.П. Скабичевским [59]установлено, что озеро Байкал
характеризуется зональностью в распределении бентосной альгофлоры. Максимальным
видовым разнообразием характеризуется литоральная зона озера, с наибольшим
диапазоном глубин обитания (0-60 м) зеленых водорослей, что связано, очевидно,
с преобладанием чистых каменистых грунтов [27].
Ведущую роль в создании первичной продукции в
литоральной зоне озера Байкал играют несколько видов макроводорослей,
образующих три первых растительных пояса: улотрикс, тетраспора,
драпарнальдиоидес. В первом поясе основным первичным продуцентом является Ulothrix
zonata (Web. et Mohr) Kütz. и
эпифитные диатомовые водоросли [30, 113].
Широко распространенный вдоль всего побережья
Байкала [30], U. zonata играет существенную роль в биогеохимических
процессах каменистой литорали озера, формировании и преобразовании среды
обитания донных гидробионтов. Заселяя обломки горных пород прибрежной зоны,
ускоряя процессы их разрушения, талломы водоросли накапливают химические
элементы, содержание которых в воде крайне незначительно, являясь, таким
образом, биогеохимическим барьером для элементов [41].
Изучение биологии и экологии водорослей рода Ulothrix
имеет важное значение в связи с тем, что они являются индикаторами пресных
чистых проточных вод [3], а сведения, связанные с особенностями
физиолого-биохимической деятельности, биологии и морфологии макрофитов, как,
например, морфометрические изменения клеток и рост культуры водорослей,
прорастание зигот, зооспор, рост гаметофита и спорофита, интенсивность
фотосинтеза и дыхания широко используют в методах биотестирования в качестве
тест-показателей [70, 144, 73, 105].
Цель работы - изучить эколого-физиологические
особенности доминирующей водоросли литоральной зоны озера Байкал U.
zonata
(Web.
et Mohr) Kütz.
Для выполнения цели были поставлены следующие
задачи:
1. Изучить особенности сезонного цикла
развития U.
zonata
на
основе наблюдений за популяцией в природных условиях и в культуре.
2. Оценить влияние U.
zonata
на биогенный и кислородный режим прибрежной зоны озера.
. Исследовать сезонную динамику биомассы
и концентрации фотосинтетических пигментов водорослей рода Ulothrix.
. На основе сравнения
морфолого-биологических характеристик вида из разных водоемов оценить
индикаторную роль U.
zonata.
Результаты работ получены в рамках комплексных
исследований лаборатории биологии водных беспозвоночных Лимнологического
института СО РАН по теме: «Междисциплинарное исследование заплесковой зоны оз.
Байкал», руководитель доктор биологических наук О.А. Тимошкин.
Полученные результаты апробированы на XVIII
Международной конференции студентов, аспирантов и молодых ученых «Ломоносов» и
опубликованы в 3-х статьях, одна из которых вышла в журнале «Известия
Иркутского государственного университета». Серия «Биология. Экология» (2011. Т.
4, № 4. С. 75-110) и две другие приняты к печати в этом журнале.
Автор признателен своим научным руководителям
д.б.н. Н.А. Бонадаренко, к.б.н., доценту Т.М. Янчук за сердечную заботу, ценные
советы и методические указания. Также выражает искреннюю благодарность д.б.н,
зав. лаб. О.А.Тимошкину за предоставленную возможность проведения работы и
консультации.
Автор благодарит д.б.н. зав. лаб. С.В. Осипову,
к.б.н. Л.А. Оболкину, к.б.н., зам. декана БПФ ИГУ по науке и НИРС А.В.
Ляпунова, к.х.н. с.н.с. В.К. Евстафьева, к.б.н., с.н.с. Н.А. Рожкову, к.б.н.,
с.н.с. О.Ю. Глызину, к.б.н., н.с. Е.П. Зайцеву, к.б.н. к.б.н. н.с. А.В.
Непокрытых, к.б.н. н.с. Н.В. Потапскую, аспирантов О.В. Попову и А.Г. Лухнева,
студентов биолого-почвенного факультета ИГУ Ю.М. Звереву и В.С. Вишнякова за
помощь при сборе материала и участие в обсуждении результатов работы.
водоросль байкал
1.
Обзор литературы
U.
zonata,
первоначально описанный как Conferna
zonata
[149], переподчинен роду Ulothrix
в первой половине XIX
века [115].
В настоящее время в соответствии сведениям об
особенностях ультраструктурной ориентации базального тела жгутикового аппарата
зооспор U.
zonata
[139, 123, 138, 117] в системе классификации водорослей данный представитель
занимает следующее положение: отдел Chlorophyta,
класс Ulvophyceae
(Lamouroux) Stewart
et Mattox,
порядок Ulotrichales
Borzi, семейство Ulotrichaceae
Kütz.
emend. Borzi,
род Ulothrix
Kütz.
[99].
U.
zonata
является одним из широко распространенных представителей своего рода, обитающим
как в пресных водоемах [135,80,
107, 105, 145, 149, 150], так и в солоноватых и морских водах [127, 92, 99,74,145,
82].
В водоемах Восточной Сибири вид отмечен: в реке
Ангара [16],в озере Байкал [17, 46, 47, 30, 34], в оз. Котакель [16], в
окрестностях города Иркутск [16], в Братском водохранилище [24, 39], в
Котельниковском термальном источнике [6], в реках Сосновка, Биракан, Таркулик,
Большая, в ручье в долине Семи озер, в оз. вблизи устья р. Давши [8], в р.
Переемная, Дулиха, Вторая Шестипалиха [33], в р. Иркут [58, 12], в р. Олха
[12], в оз. Хубсугул [16, 25], в р. Эгийн, Ценхер, Дзагастай, Марцат, Барх, оз.
Буйн [18], в р. Бальджиканка, Киркун, Кукун, Букукун, Нижний Джарлмалтай [40].
Для оз. Байкал исследователями указано на
наличие зональности в вертикальном распределении донной флоры. Первый, так
называемый, растительный пояс формирует вид U.
zonata
[17,
59, 61], дающий один пик фитомассы в период своего массового развития (конец
июня - начало июля) [30].
Появление U.
zonata
в озере отмечали в апреле - начале мая, в узкой полосе воды, образующейся у
береговой кромки льда [26]. Установлено, что в период с мая по сентябрь, в
результате повышения уровня воды в озере, граница уреза постоянно перемещается
к берегу и вслед за ней продвигается и U.
zonata
[38, 61]. Ширина образуемой им зоны (пояса) вдоль северо-западного побережья
Южного Байкала в мае - первой половине июня не превышает обычно 5 м [30]. В
конце июня - начале июля, после окончания весенней гомотермии, наступает этап
летнего прогревания воды [56, 69]. В это время отмечены максимальные (до 240
г/м2) средние величины фитомассы U.
zonata,
сплошным ковром покрывающего грунт на глубинах от 0 до 1,5 м, а ширина
растительного пояса, образованного его зарослями, достигает 15-20 м [30]. Во
второй половине июня - первой половине августа температура воды в зоне прибоя
днем в штилевую погоду может подниматься до 17-18 0С [14]. К.И. Мейер [47] указал
на то, что в этот период ярко-зеленый ровный, бархатистый ковер, образуемый
нитями U.
zonata,
изменяет свой вид: к поверхности воды тянутся многочисленные нитчатые талломы в
виде жгутов грязно-зеленого цвета длиной до 30-70 см. Автор, отметивший это
явление, связывал его с недостатком аэрации. В середине сентября с началом
устойчивого осеннего охлаждения [69], подъем уровня воды, а, следовательно,
продвижение U.
zonata
вслед
за урезом воды прекращается [14], водоросль начинает отмирать. Л.А. Ижболдина
[30] указывала, что на валунах и скалах у уреза воды узкая полоса U.
zonata
сохраняется до октября.
Наблюдения за сезонным развитием и
пространственным распределением водорослей в олиготрофных [140, 87, 130] и
эфтрофных [102, 125] озерах Северной Америки показали, что U.
zonata,
развивающийся в мелководной зоне вдоль каменистой береговой линии, имеет два
пика фитомассы: ранней весной и осенью. В летний период при повышении
температуры отмечено угнетение вегетативного развития U.
zonata
и вытеснение его другим видом нитчатых ветвящихся зеленых водорослей C.
glomerata
L. (Kütz.).
При охлаждении воды в осенний период U.
zonata
вновь возобновляет свой рост. Данные полевых наблюдений привели к формированию
гипотезы о том, что низкая температура воды и высокий уровень освещенности
играют главную роль всезонном обилии вида в северных озерах [130], а повышение
температуры более 10 0С должно сопровождаться ухудшением энергетического
баланса между фотосинтезом и дыханием водорослей [94]. При культивировании U.
zonata
в камере искусственного климата показано, что оптимальными условиями для
вегетации водоросли являются значения температуры воды 5 0С и интенсивности
освещения 1100 μЕ*m-2*s-1,
однако при этом достаточно успешно водоросли могут вегетировать при повышении
температуры воды до 15 и 20 0С. [94]. Позднее установлено, что причиной
угнетения U.zonata
и смены доминанта в мелководной зоне прибрежья североамериканских озер в летний
период является массовое бесполое размножение вида, инициируемое повышением
температуры воды [95, 120].
По продолжительности периода вегетации
водорослей в оз. Байкал Л.А. Ижболдиной [34] U.
zonata
отнесен к группе сезонных видов, вегетирующих в течение короткого периода.
Однако в конце прошлого века [30] ярко-зеленые талломы водоросли зафиксированы
в январе на глубине 0,7 м под ледяным покровом.
В озере Гурон (США) U.
zonata
также
был обнаружен под покровом льда в виде серых клочковатых обрастаний на камнях
[95]. Авторы предположили, что эти нити возникают в результате спорообразования
осенью в условиях короткого светового дня и дают начало массовому развитию
водорослей весной, которые обрастают каменистый субстрат в мелководье
литоральной зоны озера после периода ледостава.
В последнее время получены сведения о
физиолого-биохимических особенностях U.
zonata,
развивающегося в межкристаллической ледовой воде Байкала. Было показано, что
адаптация водоросли к низким температурам связана с повышением концентраций
полиненасыщенных жирных кислот (50-70%), которые играют главную роль в
поддержании текучести клеточных мембран и активности мембранно-связанных
ферментативных систем, в том числе фотосинтетической электрон-транспортной цепи
[146, 79].
U.
zonata
считается наиболее хорошо изученным представителем рода. Некоторые особенности
его морфологии, жизненного цикла и размножения были описаны еще в конце XIX
- начале XX вв. [85, 98,
131, 1,
142, 118].Во второй
половине
прошлого
столетия
появились
первые
сведения
об
особенностях
цитологии
водоросли,
знания
о
морфологии
вида
расширялись
и
дополнялись
(142, 137, 93, 141].
На основе исследований, проведенных на
культурных и природных образцах, выявлено, что вид обладает высокой степенью
полиморфизма. Сравнительные данные по морфологии и кариологии водорослей
позволили авторам прийти к заключению о необходимости слияния некоторых ранее
описанных видов рода в один - U.
zonata
[121, 142].
Вид U.
zonata
относится к категории организмов с зиготическим типом жизненного цикла [77],
когда единственной диплоидной фазой в развитии организма является зигота, образующаяся
в результате копуляции гамет, в которой затем при мейозе происходит
формирование гаплоидных спор или клеток, дающих начало новым гаплоидным
организмам [116].
Слоевище улотрикса состоит из неразвлетленных
нитей неопределенной длины, которые в начале роста прикрепляются к субстрату
базальной клеткой. Клетки нитей цилиндрические или слегка бочонкообразные,
часто короткие. Клеточные оболочки обычно тонкие, но нередко они утолщаются и
могут становиться слоистыми. Клетки содержат единственный пристенный хлоропласт
с одним или несколькими пиреноидами и одно ядро, расположенное по продольной
оси клетки. Хлоропласт имеет форму пояска, который опоясывает весь протопласт
или только часть его [22].
Размножение U.
zonata
представлено бесполым и половым путем, а также вегетативным, которое
осуществляется в результате распада нити водоросли на сегменты, развивающихся
затем в новые нити [22].
При бесполом размножении во всех клетках таллома
могут развиваться до 16 (32) четырехжгутиковых зооспор. Они бывают двух типов:
макрозооспоры, возникающие по 1-4 в клетке, и микрозооспоры, образующиеся в
большом количестве. Зооспоры различаются по форме (от эллипсоидных до
шаровидных), содержат париетальный хлоропласт, не заходящий в переднюю часть
зооспоры. Выход спор из клетки происходит через боковое отверстие в клеточной
стенке. После периода движения (имеет место положительный фототаксис) зооспора
останавливается, сбрасывая один за другим жгутики, прикрепляется боком к
субстрату и прорастает в нить [116, 7].
При половом размножении в клетках возникают
двужгутиковые гаметы в числе от 4 до 32. Гаметы выходят из клетки, заключенные
в слизистый пузырь, в котором они двигаются, пока не освободятся. Половой
процесс изогамный. У U.
zonata
наблюдается гетероталлизм. В результате копуляции образуется сначала подвижная
четырехжгутиковая зигота (планозигота), обнаруживающая отрицательный
фототаксис. После некоторого периода движения она прикрепляется к субстрату и
округляется, вытягивая один за другим жгутики. При определенных условиях зигота
прорастает в одноклеточный спорофит - Codiolum-
стадию. Сначала формируется трубковидный вырост, в который переходит содержимое
зиготы. Этот вырост позднее вздувается, а на его вершине образуются грушевидный
или дубинкообразный спорофит, содержимое которого в результате мейоза
распадается на 4-16 четырехжгутиковых зооспор. Выходя через отверстие,
образующееся в результате ослизнения части стенки спорофита, они заключены еще
в слизистую обвертку, быстро исчезающую. Зооспоры оседают на субстрат и прорастают
так же, как зооспоры, возникающие при бесполом размножении нитей U.
zonata
[116, 7].
Известно, что наиболее благоприятными условиями
для процесса спорообразования у U.
zonata,
обитающего в пресном водоеме, являются температура воды, приближенная к 20 0С,
высокий уровень освещенности, равный 520 μЕ*m-2*s-1
и фотопериод короткого (8:16 ч., день-ночь) или длинного дня (16:8 ч.,
день-ночь). При этом показано, что в условиях, когда температура воды достигает
свыше 10 0С, происходит снижение биомассы водорослей за счет повышения у них
интенсивности процесса спорообразования. Формирование гамет осуществляется в
условиях длинного светового дня (16 ч.- день) [95, 120].
В неблагоприятных для вегетации условиях талломы
U. zonata
переходят в пальмелевидное состояние. Этому предшествует утолщение и ослизнение
оболочек клеток, что служит защитой от эпифитов и повышает устойчивость клеток
в колонии [76]. В.М. Арнольди [1] объяснял такую особенность водоросли, как
специфическую реакцию на недостаток в воде растворенного кислорода.
2.
Объекты и методы исследования
Сбор полевого материала.
Сбор макроводорослей на глубине от 0 до 1,5-3 м осуществляли с использованием
щипцов Рубцова. С помощью скальпеля отделяли водорослей от камней. Пробу
промывали через капроновое сито, переносили в емкость для хранения и
фиксировали раствором формалина до 4% конечной концентрации [45].
Для подготовки материала к проведению
молекулярно-биологического анализа пробы отбирали в бухте Большие Коты, заливе
Листвяничный, проливе Малое море, на острове Большой Ушканий, о-ве Ольхон.
Образцы фиксировали спиртом (96%), а также после удаления излишков воды
помещали в полипропиленовые пробирки типа эппендорфа объемом 1,5 мл с
силикагелем.
В работе используются данные изменений
химического состава воды в прибрежье озера, предоставленные д.г.н., с.н.с. В.М.
Домышевой и к.б.н. М. В. Сакирко, а также - неопубликованные данные о динамике
биомассы микроводорослей в толще водыд.б.н. Н.А. Бондаренко.
Спектрофотометрический анализ
пигментов. В задачи данного исследования входило изучение
сезонной динамики фитомассы основной водоросли урезовой зоны оз. Байкал Ulothrix
zonata
Kütz.,
содержания пигментов в её клетках и соотношения между этими показателями.
Спектрофотометрический анализ пигментов U.
zonata
проходил на базе СИФБР СО РАН под руководством д.б.н. С.В. Осиповой. Для
изучения концентраций пигментов материал отбирали в период открытой воды в
западной части Южного Байкала в зал. Листвяничный и в бух. Большие Коты с июня
по октябрь 2011 г. на расстоянии от 0,2 до 4 м от уреза воды с глубин от 0,3 до
0,5 м, во время ледостава - в заливе Листвяничный в феврале и марте 2012 г. под
ледяным покровом, окаймляющим берег озера, - заберегом, с помощью щипцов
Рубцова. В качестве руководства для отбора проб, их хранении, проведении
анализа и обработке результатов использовался стандарт ГОСТ 17.1.4.02-90 «Вода.
Методика спектрофотометрического определения хлорофилла а».
Перед проведением анализа предварительно
водоросли очищали от загрязнений и промывали дистиллированной водой. Излишки
воды удаляли фильтровальной бумагой. Навески гомогенизировали в фарфоровых
ступках с добавлением ацетона (90%) и кварцевого песка в течение 15 минут.
Светорассеивающую взвесь удаляли из экстракта центрифугированием на аппарате K26D
(Германия) с ускорением 8000 оборотов/мин. в течение 15 минут. После
центрифугирования экстракт доводили до объема фотометрической кюветы. При
спектрофотометрировании использовали кюветы с рабочей длиной 1 см. Отсчеты
оптических плотностей брали на шести длинах волн- 430, 480, 664, 647, 630 и 750
нм. Фотометрирование проводили дважды: до и после подкисления экстракта
несколькими каплями приготовленного раствора соляной кислоты в ацетоне.
При обработке данных использовались стандартные
программные пакеты для персонального компьютера.
Метод получения культур.
Собранный в природе живой материал служил источником получения лабораторной
культуры водорослей. Культивирование проводили в Центре коллективного
пользования «Пресноводный аквариумный комплекс» (ЦКП ПАК). Для этого
использовали аквариальную установку. В аквариум, наполненный байкальской водой,
помещали камни с обрастаниями. Температура воды составляла £
4 °
С. Температура воздуха не превышала 7,5° С. В качестве
источника освещения использовали люминесцентные лампы общего назначения.
Небольшой фонтан моделировал волновое движение воды в аквариуме.
В условиях экспериментальной лаборатории
исследовали морфологические и биологические особенности U.
zonata
с сентября 2010 года по июнь 2012 г. В ходе эксперимента дважды в неделю при
микроскопировании наблюдали состояние культивируемой водоросли, фотографировали
и делали замеры клеток.
Материал, полученный в условиях лаборатории,
помещали в чашки Петри на северное окно лаборатории. Периодически (по мере
подсыхания) в чашки добавляли воду из исходного местообитания и питательную
среду Z-8 [133], имеющую
активную реакцию (рН), как в исходной среде. Такие смешанные культуры служили
источником живого материала.
Для выделения нитей U.
zonata
использовали
пипеточный метод: при малых увеличениях светового микроскопа (×
10-20) с
помощью стерильной пипетки Пастера с тонко оттянутым длинным концом отлавливали
единичные клетки или нити. Водоросли, отловленные благодаря всасывающей силе
капилляра, переносили из одной капли стерильного питательного раствора в
другую, пока в капле не оставалась искомая водоросль с максимальным отсутствием
посторонних видов, после чего чашки Петри выставляли на естественный рассеянный
свет при комнатной температуре [12].
Обработка материала.
Идентификацию, изучение особенностей морфологии водорослей проводили с
использованием светового микроскопа MEIJI
TECHNO CO.
LTD. при увеличениях ×100
и
×400.
Результаты
исследований документировали с использованием фотоаппарата Olympus
C-3040 zoom
(3.3Mgpxl) с
фотонасадкой NY 2000S
01705.
Для определения биомассы использовали весы Iuchi
Sefi (IB-200H) (Япония) и Adventurer
Ohaus AR
2140 (Китай).
Для сканирующей электронной микроскопии (СЭМ)
материал дегидратировали в растворах этилового спирта возрастающей
концентрации. Полное высушивание препаратов проводили при температуре 40º
С,
после чего наносили на столики для СЭМ и напыляли золотом (напыление
проводилось в приборе SCD-004 (Balzers). Столики с образцами исследовали в
сканирующем электронном микроскопе FEI Company Quanta 200 в режиме высокого
вакуума.
Экспресс-оценка влияния
нефтезагрязнения. Оценку влияния загрязнения водоема
нефтепродуктами на морфометрические показатели U.
zonata
проводили
на основе сравнения популяций из р. Ангара, р. Олха и оз. Байкал. Во всех трех
водоемах камни с обрастаниями брали на глубине примерно 0,5 м. При
микроскопировании исследовали особенности морфологии клеток в талломах.
Методика учета тератогенных особей заключалась в следующем: в 200 случайных
полях зрений считали общее количество клеток и количество аномальных клеток,
определяли их процентное соотношение.
Статистическая обработка
экспериментальных данных. При статистической обработке
экспериментальных выборок использовали стандартные компьютерные программы
("МатЛаб", "Статистика", а также встроенные в "Excel"
и графический редактор "Graf4Win").
Высчитывались среднее арифметическое значение и несмещённое среднеквадратичное
отклонение, которые использовались как оценочные для матожидания и дисперсии
генеральной совокупности [5]. Далее устанавливалось, какому из канонических
распределений в наибольшей степени соответствует данная выборка. Доверительный
интервал вычислялся, исходя из 95%-ого уровня доверия, по формуле
d = ± 1.96sn-1,
где sn-1
- несмещённое среднеквадратичное отклонение.
Ошибка среднего значения находилась по формуле
[5, стр. 36-37]:
где d0 - ошибка
единичного измерения ,
dn - ошибка среднего
серии измерений,
n - число измерений
(объём выборки).
3.
Результаты и обсуждение
.1
Особенности сезонного цикла развития доминирующего вида макроводорослей в
прибрежной зоне оз. Байкал U.
Zonata
В ходе полевых наблюдений в зимний период в
районе истока р. Ангара обнаружены нитчатые талломы водорослей ярко-зеленого
цвета, растущие на небольших валунах и камнях под ледяным покровом, окаймляющим
берег озера, - заберегом. Ширина ледяного покрова в феврале и марте составляет
примерно 5 м, толщина льда- 30 см, глубина воды в прибрежной зоне, покрытой
льдом - 0,5-0,7 м. И работах других исследователей зимняя популяция U.
zonata
в оз. Байкал была зафиксирована только в период ледостава, когда водная
поверхность полностью покрывается льдом.
Выявлено, что обрастания каменистого прибрежья,
развивающиеся под первичным заберегом, в феврале и марте представлены, главным
образом, водорослями родов Ulothrix,
Spirogyra, Stigeoclonium,
Didymosphenia, Stratonostoc.,
Cladophora.
В пробах, собранных с нижней поверхности
заберега,
нити U.
zonata
всегда встречаются совместно с зеленой нитчатой водорослью Spirogyra
sp. Elenk. Обилие последней в
феврале значительнее. Талломы U.
zonata,
представленные преимущественно в виде фрагментов разной длины, состоящих из
клеток с поясковидным хлоропластом, конечные клетки часто имеют закругленную
внешнюю сторону (рис. 1). Эти морфологические особенности вида являются
признаками интенсивного вегетативного деления, в результате которого, вероятно,
пространственное распределение осуществляется более эффективно. Средние
значения ширины клеток U.
zonata
в
феврале составляли 21,2 км,длины - 10,5. Эпифитирующими на талломах улотрикса
отмечены колонии Didymoshenia
geminata
(Lyngb.)
M. Schmidt.,
Didymosphenia
dentata
(Dorogostaisky)
Skv., Didymosphenia
sp., видов родов Nitzschia
Hass. и Navicula
Bory.
В марте в некоторых подледных участках отмечены
многочисленные нити U.
zonata,
которые, поднимаясь от камней к нижней поверхности льда, образуют в толще воды
своеобразные зеленые столбы длиной до 50-70 см и, порой, врастают в лед.
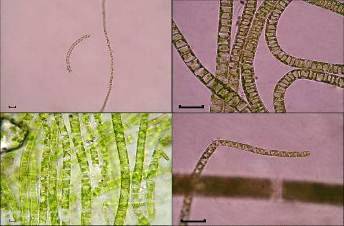
Рис. 1. U.
zonata,
вегетирующие в феврале под ледяным покровом, окаймляющим берег озера
Формируя ярко-бархатистый ковер, расстилающийся
по каменистому дну и вдоль внутреннего края заберега (рис. 2), не только подо
льдом, U.
zonata
является
доминантным видом в этой зоне озера, обилие талломов других нитчатых водорослей
ему уступает.

Рис. 2. Обрастания U.
zonata,
формируемые на каменистом субстрате вдоль береговых ледовых наростов
При микроскопировании у U.
zonata
отмечен процесс массового спорообразования и выход зооспор (рис. 3).
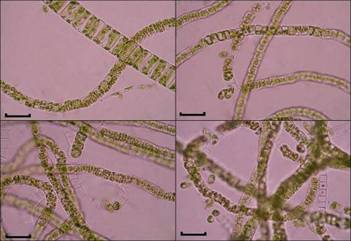
Рис. 3.U.
zonata,
вегетирующий в марте под ледяным покровом, окаймляющим берег озера
Как следует из рисунка (рис. 5), средние
значения для ширины и длины клеток U.
zonata
в марте - 22 и 12 мкм соответственно. Эти значения так же, как и в феврале,
ниже средних значений размеров клеток в летний период. Однако фитомасса,
средняя величина которой в марте составляет 202,7 г/м2, близка к таковой в
период, отмечаемый, как летний пик развития вида в Южном Байкале и равна 210, 6
г/м2. Это позволяет сделать предварительный вывод о том, что в зимнее время
наблюдается второй пик развития U.
zonata
в
мелководье литоральной зоны озера и выделить две популяции водоросли: зимнюю и
летнюю.
Данные о первичной структуры области второго
внутреннего транскрибируемого спейсера кластера рибосомных генов (ITS2)
указывают на существование 1% внутривидовой изменчивости между летней и зимней
популяциями U.
zonata
в оз. Байкал (данные, полученные совместно с доктором Кристианом Боэдекером,
Новая Зеландия, подготовлена к печати статья).
В апреле в зоне уреза воды, первой
освобождающейся ото льда, на каменистом субстрате отмечаются обрастания U.
zonata
в
виде молодых вегетативных нитей. Средние значения ширины клеток - 19,5 мкм,
длины- 12,3 мкм. После освобождения прибрежья от заберегов, в талой воде на
расстоянии 1 - 2,5 - 3 м еще можно наблюдать зимнюю популяцию, пик вегетации
которой приходится на февраль - март. В этой зоне обрастания имеют темно-зеленый
цвет, отчасти из-за присутствия большого количества песка, вероятно, попавшего
в заросли водорослей при весеннем перемешивании воды в озере. Талломы U.
zonata
представляют собой короткие нити, разрушавшиеся в тех частях, где располагались
многочисленные опорожнившиеся спорангии (рис. 4).
Не смотря на то, что в некоторых нитях все еще
наблюдается процесс спорообразования, выход зооспор или их прорастание из
материнской клетки, утолщение оболочек клеток, разрушение хлоропласта, обильное
обрастание эпифитами и, в некоторых случаях, формирование специфических
структур, похожих на пальмелевидное состояние, указывают на то, что водоросль
находится на стадии отмирания. Средние значения размеров клеток водоросли в
этот период указаны на рисунке 5.
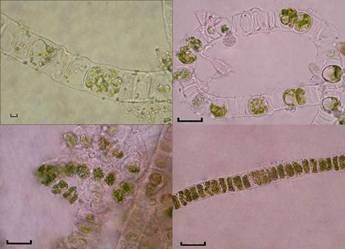
Рис. 4. U.
zonata.
Завершение вегетации зимней популяции
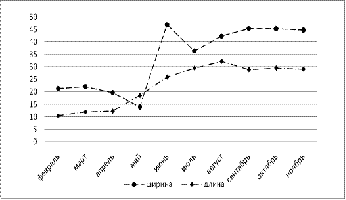
Рис. 5.Сезонная динамика морфометрических
показателей вида.
В июне-июле наблюдается летний пик развития вида
[30]. Средние значенияширины клеток 46,8 и 36,3 мкм, длины- 25,8 и 29,4 мкм
соответственно. В летний максимум фитомассы зафиксировано активное вегетативное
деление нитей, а также процесс массового спорообразования (рис. 6).

Рис. 6. U.
zonata
в летний пик развития
В штилевую погоду вода в прибрежье озера
приобретает светло-зеленый цвет. Это явление обусловлено массовым выходом
четырехжгутиковых зооспор U.
zonata.
Их концентрация в толще воды представлена значением 1,77 х 105 кл/мл или
177 тыс. кл/мл. С помощью СЭМ указаны точные размеры клеток бесполого
размножения U.
zonata:
длина спор равна 5,6 мкм (SD=0,7),
ширина - 4,4 мкм (SD=0,8)
(рис.7).
Возрастная структура популяции не однородна в
течение всего периода вегетации. В июле и августе наблюдается уменьшение
средних значений ширины клеток, и, наоборот, увеличение длины (см. рис. 5). Это
связано с тем, что доля молодых особей в популяции возрастает вследствие
активного бесполого размножения в июне.
Рис.7. Зооспоры U.
zonata
(СЭМ)
На крупных валунах и скалах у уреза воды нити U.
zonata можно встретить до ноября [34]. Нами отмечено, что в условиях
экспериментальной установки вегетация водорослей продолжается до декабря. В это
время средние значения ширины клеток составляет 45,12мкм, длины- 29,08 мкм.
В ходе изучения особенностей сезонного развития U.
zonata нами не удалось проследить выход гамет и их копуляцию. Однако в
осенний период, когда вегетация водорослей завершается, в прибрежье оз. Байкал
и в культуре при микроскопировании отмечены многочисленные клетки округлой и
дубинкообразной формы с утолщенной оболочкой и плотным, зернистым содержимым
ярко-зеленого цвета, - одноклеточные спорофиты (Codiolum-
стадия), формирующиеся в результате прорастания зиготы (рис.8). Это дает
основание предположить, что споры, выходящие из спорофита, дают начало
популяции U. zonata, вегетирующей в озере в период ледостава и дающей
второй пик фитомассы в марте.
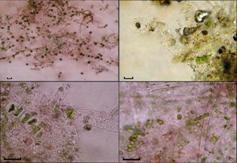
Рис. 8. Одноклеточные спорофиты - Codiolum-
стадия
Терминальные клетки таких нитей не
дифференцированы на апикальную и базальную, их внешняя сторона имеет
закругленную, обтекаемую форму. Мы предполагали, что такая морфологическая
структура талломов U.
zonata,
по типу планктонных диатомовых водорослей родов Aulacoseira,
Melosira ориентирована на
передвижение в толще воды. Появляясь в осенний период, эти нити предназначены
для миграции из прибрежной зоны, покрываемой льдом и полностью промерзающей в
зоне уреза, вглубь озера, где в дальнейшем под покровом льда являются
источником «ледовой» популяции, сведения о которой появились недавно [146, 79].
Однако результаты молекулярно-биологического анализа (данные получены в
совместных работах с доктором Кристианом Боэдекером , Новая Зеландия) показали,
что встреченные нами нити относятся к другому роду, ранее не отмеченному для
оз. Байкал. Для точной идентификации вида необходимы сведения
ультраструктурного анализа клеток водорослей.
3.2
Сезонные изменения биомассы и концентраций фотосинтетических пигментов и U.
Zonata
Ведущая роль в функционировании пресноводных
экосистем принадлежит макро - и микроводорослям, за счет фотосинтеза которых в
водоемах создается фонд органического вещества, составляющий энергетическую
основу для всех последующих этапов продукционного процесса в водоеме [10]. К
одним из наиболее распространенных показателей, используемых при изучении
водорослевых сообществ, относятся фотосинтетические пигменты [49, 50].
Содержание основного пигмента зеленых растений,
хлорофилла а, считается универсальной эколого-физиологической
характеристикой развития и фотосинтетической активности водорослей [49]. Кроме
того, в отечественной литературе часто используются такие пигментные
характеристики альгоценозов, как концентрация хлорофиллов в и с,
каротиноидов, пигментные индексы [9, 19, 21, 44, 53, 56, 61]. Колебания
значений этих пигментных показателей принято объяснять соответствующими
изменениями в составе, обилии или физиологического состояния альгосообществ [9,
21, 49, 55].
В настоящее время все чаще ставят знак равенства
между хлорофиллом и биомассой, которую выражают в концентрации пигмента.
Преимущество хлорофилльного метода, по сравнению с микроскопическим учетом
водорослей, заключается в скорости получения результатов, так как часто
возникает необходимость оперативной оценки временной и пространственной
динамики альгоценозов в разных экологических условиях, а также в ходе сезонной
или многолетней сукцессии [51].
Известно, что удельное содержание хлорофилла a
в сырой биомассе водорослей значительно варьирует в зависимости от многих
факторов: светового режима [80, 126,42,
87], сезона года [70, 42] обеспеченности водорослей минеральным питанием [70,
61], температурных условий [42, 87], времени суток [120] и др., что не
позволяет вывести общий переходный коэффициент между концентрацией хлорофилла и
биомассой, пригодный для водоемов разного типа [20, 62, 130,124].
В научной литературе зависимость содержания
хлорофилла a от биомассы фитопланктона широко обсуждается для разных
водных экосистем [19, 35, 86, 81, 87, 94, 105, 110,145, 146]. В подобных
исследованиях наблюдается как линейная [62,145], так и нелинейная связь
хлорофилла и общей биомассы водорослей. Значительно варьируют и получаемые
коэффициенты корреляции между данными параметрами. [44, 87, 110, 146].
Для фитопланктона оз. Байкал такая зависимость
прослежена [50]. Однако для макрофитобентоса, являющимся основными продуцентом
органического вещества в литоральной зоне озера, приведены единичные данные о
биомассе и фотосинтетической активности [29, 30,113] при этом связь хлорофилл-биомасса
макроводорослей изучена лишь в кратковременном аспекте [113]. В то время как,
считается, что переходный коэффициент между данными параметрами может быть
получен лишь при многолетних наблюдениях [61, 62].
Одной из задач настоящей работы являлось изучение
сезонной динамики фитомассы и концентраций основных фотосинтетических пигментов
доминирующей водоросли в прибрежной зоне оз. Байкал U.zonata.
Общая динамика накопления основных
фотосинтетических пигментов (хлорофиллов а и в, каротиноидов) и
биомассы в сезонном цикле развития вида представлена на рисунке 8.
Исследование показало, что максимальные значения
биомассы приходятся на июль (210,6 г/м2) и март (202,3 г/м2) - летний и зимний
пики развития вида в прибрежье. В это же время отмечены повышенные, но не
максимальные, концентрации хлорофилла а в клетках водорослей, которые
составляют 17 и 33 мг/м2 соответственно. Корреляционный анализ усредненных
данных выявил наличие тесной и высоко значимой положительной корреляции (r=0,79)
между этими показателями для летней популяции (июнь-октябрь).
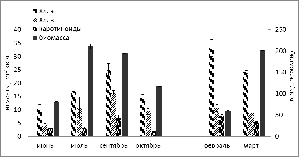
Рис. 8. Сезонная динамика концентраций пигментов
(хлорофилла а (Хл.а) и в (Хл.в), каротиноидов)
(мг/м2) и биомассы U.
zonata
(г/м2).
Зависимость между накоплением биомассы и содержанием
хлорофилла а у зимней популяции (февраль-март) принимает отрицательное
значение (r=-1). Как
указывалось выше, удельное содержание хлорофилла в биомассе водорослей
значительно варьирует в зависимости от многих факторов. Хорошо известно, что
интенсивность света контролирует содержание хлорофилла у высших растений [55].
У водорослей свет также является одним из факторов, определяющим концентрации
пигментов в клетках, как в природных сообществах [70, 81], так и в условиях
культуры [126].Возможно, увеличение содержания клеточного хлорофилла у U.
zonata,
регистрируемое в феврале на фоне сравнительно невысоких значений биомассы,
может происходить в ответ на лимитированные световые условия, так как в это
время камни с обрастаниями можно наблюдать преимущественно под покровом
заберегов.
Максимальная концентрация хлорофилла а (24,8
мг/м2), а также вспомогательного фотосинтетического пигмента зеленых водорослей
хлорофилла в (16,3 мг/м2) зафиксированы в сентябре. В это время
обрастания каменистой береговой линии все еще представлены вегетирующими
талломами, формирующими на поверхности субстрата густые ватообразные скопления
темно-зеленого цвета. Этим можно объяснить высокие значения биомассы и
концентраций основных фотосинтетических пигментов, тесную связь которых отражает
высокий коэффициент корреляции (r=
0,79 и r=0,80).
В это время, а так же в период, когда развитие
водорослей происходит под ледяным покровом, окаймляющим берег озера,
зарегистрированы максимальные значения содержания каротиноидов в клетках U.
zonata
-
6,9 и 7,6 мг/м2 соответственно. Известно, что преобладание концентраций желтых
пигментов является признаком неблагополучия водорослей, связанного со старением
сообщества, истощением минерального питания, недостатком или избытком
освещенности [89, 61, 70]. Повышение концентраций этих пигментов в сентябре
может быть связано с возрастанием доли «стареющих» особей в популяции или же
уменьшением длины светового дня. Последнее может является сигналом для
прекращения вегетации водорослей и, как следствие, снижения биомассы в октябре.
Для летнего периода развития U.
zonata
прослежена
положительная корреляционная связь между изменениями биомассы и концентраций
каротиноидов (r=0,55). Зависимость
концентраций желтых пигментов от изменения биомассы в период промерзания
береговой линии не выявлена (r=-1),
что может быть обусловлено сменой фактора, определяющего содержание пигментов.
Сезонная динамика биомассы и концентраций
основного фотосинтетического пигмента хлорофилла а характеризуется
минимальными значениями в июне и октябре, когда популяция в первом случае
преимущественно представлена молодыми талломами водорослей и завершающими
вегетировать - во втором.
Приведенный анализ данных по содержанию
пигментов и биомассы U.
zonata
в периоды ледостава и открытой воды в озере показал наличие тесных прямых
корреляций между этими показателями только в летний период развития вида.
3.3
U.
Zonata
как средообразующий вид прибрежной зоны озера
Байкал
В ходе проведенных исследований выяснено, U.
zonata оказывает влияние на кислородный и биогенный режим прибрежья озера.
Сезонные изменения рН, концентраций кислорода и биогенных элементов в прибрежье
озера в период июнь-сентябрь представлены на рисунке 9.
Рис.9. Динамика рН, концентраций кислорода и
биогенных элементов в прибрежье озера в период июнь-сентябрь 2010 года (по:
Тимошкин и др., 2012, данные В.М. Домышевой и М.В. Сакирко)
Как показал химический анализ воды наибольшие
значения концентраций кислорода (13,23 мг/л) и ионов водорода (pH=9),
а также незначительная величина концентраций ионов гидрокарбонатов (62,7 мг/л)
приходится на июль, когда наблюдается летний пик развития вида. Затем значения
этих параметров изменяются: концентрация кислорода и ионов водорода - в стону
уменьшения, концентрация ионов гидрокарбонатов - увеличения, что может
свидетельствовать о снижении интенсивности процесса фотосинтеза водорослей с
наступлением осеннего периода.
Поскольку U. zonata - доминирующий вид на
глубине от 0 до 1,5 м, то он является основным потребителем биогенных элементов
в этой зоне, в частности фосфатов и нитратов. Изменения концентраций этих
веществ в прибрежной воде могут изменяться в зависимости от преобладающей фазы
жизненного цикла водоросли в том или ином месяце.
В период летнего пика развития U. zonata,
в июле, отмечены минимальные концентрации нитратов в воде (0,0294 мг/л) (см.
рис. 9), а так как азот входит в состав важнейших белков, нуклеиновых кислот,
аминокислот, хлорофилла, ферментов, многих витаминов, липоидов и других
органических соединений, он является одним из основных элементов, необходимых для
интенсивного роста растений.
К середине сентября значительно уменьшалось
количество соединений фосфора в воде (см. рис. 9). Фосфор входит в состав
сложных белков (нуклеопротеидов), нуклеиновых кислот, фосфатидов, ферментов,
витаминов, фитина и других биологически активных веществ, участвует в обмене
веществ, делении клеток, размножении, передаче наследственных свойств. В этом
месяце в талломах водорослей наблюдали интенсивное формирование
продуктовразмножения.
Поскольку биомасса водорослей в толще воды в исследуемый
период была крайне мала,- от 27 до 255 мг/м2 (рис.10.), а обрастания на
талломахU. zonata составляли от 0,1 до 11 % (табл.1), то основным
фактором, влияющим на эти изменения являлась фитомасса U. zonata,
максимальные значения которой зарегистрированы в июле и составили210,6 г/м2
(см. рис.8).
Таблица № 1. Процентное соотношение
микроводорослей, эпифитирующих на талломах U.
zonata
в летний период (июнь-август)
|
Месяц
|
Встречаемые
водоросли, %
|
|
Hannaea
sp.
|
Gomphonema
sp.
|
Cymbella
sp.
|
Nitzschia
sp.
|
Navicula
sp.
|
Cocconeis
placentula
|
|
Июнь
|
5,6
|
6
|
0
|
0
|
0
|
|
Июль
|
0,3
|
0,04
|
0
|
0
|
0,9
|
0
|
|
Август
|
11,3
|
7,6
|
0,7
|
5,9
|
11,4
|
0,1
|
Ранее было установлено, что U.
zonata
является не только основным источником первичной продукции в прибрежной зоне
оз. Байкал, но и пищей и убежищем для донных беспозвоночных [34, 38; 52.
53].
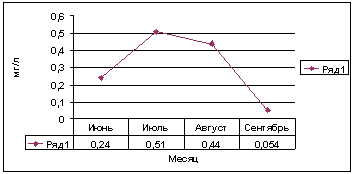
Рис. 10. Динамика биомассы микроводорослей в
толще воды (по: Бондаренко и др., 2012).
Наши исследования показали, что U.
zonata
влияет на кислородный и биогенный режим прибрежья и, как следствие, является
видом - эдификатором в этой зоне озера.
3.4
Экспресс-оценка влияния загрязнения водоема нефтепродуктами на водоросль u.
Zonata
на основе сравнения морфолого-биологических параметров популяции из р. ангара,
р.Олха и оз. Байкал
Водная экосистема понимается как единство среды
и обитающей в ней биоты. Она формируется под действием и в результате
процессов, протекающих в бассейне водосбора и на протяжении всего русла реки.
Химический анализ, поэлементно оценивая среду обитания, лишь косвенно может
указывать факторы, оказывающие влияние на экосистему или являющиеся результатом
ее жизнедеятельности [4].
Действие токсических веществ, оказывающих
вредное воздействие на организмы, по мере их влияния на экосистемы можно
разделить на фазы острой и хронической токсичности. Для определения острой
токсичности служат экспресс-методы продолжительностью в несколько дней (один -
три и более), а для определения хронической токсичности - продолжительные опыты
(месяц и более) [63].
Наиболее адекватно состояние водной экосистемы
можно оценивать по составу сообществ водных организмов [2]. Причем в ряде
отечественных и зарубежных систем оценки используются показатели и индексы,
связанные с развитием той или иной группы организмов от рыб до водорослей [64].
Биоиндикационные методы на основе оценки
водорослевых сообществ дают интегральную оценку результатов всех природных и
антропогенных процессов, протекавших в водном объекте. Кроме того, биоиндикация
по сообществам водорослей является бюджетным экспресс-методом, в то время как
проведение необходимых химических анализов зачастую оказывается весьма
дорогостоящим [4].
Преимуществом автотрофов является то, что они
первыми в трофической цепи реагируют на загрязнители, не успевая их значительно
накапливать [4].
Зеленые водоросли часто используются в методах
биотестирования, причем как целые сообщества, так и отдельные виды [65, 63].
Известно, что данные, связанные с особенностями биологии макрофитов, как,
например, морфометрические изменения клеток, используются в качестве
тест-показателей, изменяющимися под действием неблагоприятных факторов [98,
142, 71, 103].
U.
zonata
обитает в пресных чистых проточных водоемах и считается индикатором таковых
[3], в связи с этим представляло интерес оценить его индикаторную функцию в
ответ на влияние загрязнения водной экосистемы нефтепродуктами на основе
сравнения морфолого-биологических параметров популяций из разных водоемов.
апреля 2012 года в результате повреждения
нефтепровода ФГУ Комбината «Прибайкалье» Росрезерва произошла утечка дизельного
топлива в р. Ангара. По сообщению пресс-службы правительства Иркутской области
содержание нефтепродуктов у оголовка водозабора превышала допустимое значение
0,3 мг/л и составляло 23 мг/л. В качестве представителей экспертной организации
мониторинга выступал филиал Центра лабораторного анализа и технических
измерений по Восточносибирскому региону. По данным экспертов управления, объем
нефтепродуктов, поступивших в реку, составил не менее 316 тонн. Часть
нефтепродуктов с помощью сорбентов была собрана ниже по течению. В районе, где
произошла авария, скорость течения реки составляет примерно 1 м/с, постепенно
снижаясь к подпору Братского водохранилища (г. Свирск). 6 мая пятно на
поверхности реки наблюдалось на расстоянии 180-200 км от места аварии. Отбор
проб обрастаний U.
zonata
проводился в этом районе.
Сравнительный морфолого-биологический анализ
показал, что в нитчатых талломах водорослей, отобранных в прибрежье р. Ангара
довольно часто встречаются клетки по своим морфометрическим особенностям не
характерные для данного вида в нормальных условиях. Отмечены различные вздутия,
неравномерные утолщения клеточных стенок, формирование своеобразных выростов, в
которые словно перетекало содержимое соседних клеток. Аномальные
морфологические признаки зарегистрированы не только на уровне отдельных клеток,
но, хоть и в редких случаях, на уровне нитей, когда, к примеру, таллом
водоросли в определенном участке нити напоминает скрученный зеленый канат (рис.
11).
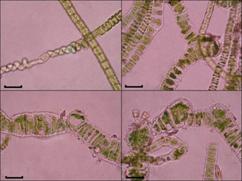
Рис.11. Аномальные клетки в талломах U.
zonata
В такой структуре сложно различить отдельные
клетки и их хлоропласты. Кроме того, при микроскопировании также отмечали
колонии различной формы и размера, покрытые слизью, которые обычно формируются
в результате неправильно поделившихся, во всех направлениях, клеток -
пальмелевидное состояние. Известно, что подобное происходит, когда U.
zonata
оказывается в неблагоприятных условиях, связанных с недостатком растворенного
кислорода (Boney, 1981; Арнольди,1925)
В р. Ангара выявлен значительный процент аномальных
клеток внутри талломов водорослей (до 1,9 %), в то время как в р. Олха и оз.
Байкал тератогенные особи не регистрировались, что является доказательством
чистоты воды двух последних экосистем.
Полученные данные свидетельствуют о том, что
водоросли вида U.
zonata
могут использоваться в методах экспресс-оценки влияния загрязнения
нефтепродуктами водных экосистем на их компоненты.
Выводы
Изучение материала, полученного в ходе натурных
наблюдений и культурального исследования с применением эколого-физиологических
и молекулярно-биологических методов, позволяет сделать следующие выводы:
. В урезовой зоне прибрежья южной котловины
озера Байкал, Малого Моря, Ушканьих островов обитает Ulothrix
zonata,
а вид U.
tenuissima
Kütz.,
приводимый ранее в списках исследователей макроводорослей озера (Ижболдина,
2007), не выявлен.
. Выявлены особенности биологии U.
zonata,
обитающего в озере Байкал. U.
zonata
не
является видом короткого периода вегетации, что предполагалось исследователями
ранее: одну (или несколько одновременно) из его жизненных стадий можно
обнаружить в урезовой зоне Байкала в течение круглого года. Представлены
фотодокументы каждой стадии развития.
. Впервые в цикле развития описан зимний период
(январь-март) и доказано, что в Байкале вид формирует две популяции: зимнюю и
летнюю. В зимний период (январь-март) в районе истока р. Ангара обнаружены
нитчатые талломы водорослей, растущие на небольших валунах и камнях под ледяным
покровом, окаймляющим берег озера (заберегом), где U.
zonata,
как и в летний период, формирует первый растительный пояс. Сведения по биомассе
вида в летнее и зимнее время позволяют выделить два пика развития U.
zonata
в прибрежье озера.
. Анализ данных о химическом составе воды и
изучение сезонной динамики фитомассы U.
zonata
показали, что улотрикс не только является основным видом, создающим биомассу
прибрежья, но и влияет на кислородный и биогенный режим этой зоны озера.
. Приведены первые данные об особенностях
динамики концентраций основных фотосинтетических пигментов (хлорофиллов а, в
и каротиноидов) и биомассы U.
zonata
в
сезонном аспекте. Изучение динамики содержания основных пигментов, служащих
универсальной эколого-физиологической характеристикой развития и
фотосинтетической активности водорослей, показало, что колебания значений этих
пигментных показателей соответствовало изменениям в обилии или физиологическом
состоянии как зимней, так и летней популяций водоросли.
. Результаты исследования морфологических
изменений клеток U.
zonata,
роста культуры, наблюдения за прорастанием зигот, зооспор, ростом спорофита
представляют интерес не только для теоретических работ в области экологии и
биологии водорослей, но также могут быть использованы при проведении
экологического мониторинга водных экосистем, что продемонстрировано нами при
исследовании популяций улотрикса не только из озера Байкал, реки Олха, но и из
загрязненного нефтепродуктами района реки Ангара. В последнем биотопе внутри
талломов водоросли нами выявлен значительный процент аномальных клеток (до 1,9
%), в то время как в чистых экосистемах (р. Олха и оз. Байкал) тератогенные
особи не регистрировались. Этот анализ позволяет использовать данный вид в
методах экспресс-оценки влияния загрязнения нефтепродуктами водных экосистем.
Список
литературы
1. Арнольди В. М., Введение в
изучение низших организмов. Морфология и систематика зеленых водорослей и
близких к ним окрашенных орагнизмов., М., 1901, [3 изд.], М.-Л., 1925.
. Баринова С. С. Полиморфизм
соединительных структур диатомовых водорослей // Эволюционные исследования.
Вавиловские темы. Владивосток, 1988. С. 110-122.
. Баринова С.С., Медведева
Л.А., Анисимова О.В. 2000. Водоросли-индикаторы в оценке качества окружающей
среды // М.: ВНИИ природы. 150 с.
. Баринова С.С., Медведева
Л.А., Анисимова О.В. 2006. Биоразнообразие водорослей-индикаторов окружающей
среды // Тель-Авив: Инст. Эволюции Университета Хайфы. 498 с.
. Бейли Н. Статистические
методы в биологии. М.: ИЛ, 1962. 260 с. ИБВВ № 7626.
. Биота некоторых термальных
источников Прибайкалья и связанных с ними водоемов/ Тахтеев В.В. [и др.] //
Исследования водных экосистем Восточной Сибири. - Иркутск: Изд-во Иркут. ун-та,
2000. - С. 55-100.
. Ботаника. Курс альгологии и
микологии. Учебник / Под ред. Ю.Т. Дьякова. - М.: Изд-во МГУ, 2007. - 559 с.
. Бочка А.Б. Водоросли //
Флора и фауна заповедниов. Флора и фауна водоемов и водотоков Баргузинского
заповедника. - М., 2000.- С. 8-123.
. Бульон В.В. (1983)
Первичная продукция планктона внутренних водоёмов. Л., Наука, 150 с.
. Винберг Г. Г. Первичная
продукция водоемов. - Минск: Изд-во АН БССР, 1960. - 328 с.
. Вишняков В.С. Новые
сведения о макроводорослях залива Большие Коты и его притоков (Южный Байкал,
Россия) // Актуальные проблемы современной альгологии: Тез. докл. IV
Международной конф. (г. Киев, 23-25 мая 2012 г.). - Киев, 2012а. - с. 55-56.
. Вишняков В.С. К
характеристике альгофлоры водоемов долины нижнего течения реки Иркут (Восточная
Сибирь, Россия) // Актуальные проблемы современной альгологии: Тез. докл. IV
Международной конф. (г. Киев, 23-25 мая 2012 г.). - Киев, 2012б. - с. 57-58.
. Водоросли: Справочник //
Под общ. ред. С.П. Вассера.-Киев: Наук, думка, 1989.-608 с.
. Вотинцев К.К. Гидрохимия
озера Байкал. Тр. Байкальского лимнологического института. Издательство АН
СССР, т ХХ., 1961.
. Галазий Г.И., Картушин
В.М., Лут. Б.Ф. Атлас Байкал, Москва, 1993.- С. 160.
. Дорогостайский В.Ч.
Материалы по альгологии оз. Байкал и его бассейна // Изв. Вост.-Сиб. отд. Русс.
геогр. об-ва. 1904. Т.35, вып. 3.
. Дорогостайский В.Ч.
Материалы для альгологии оз. Байкал и его бассейна // Изв. Вост.- Сиб. отд.
Император. Рус. геогр. О-ва. - 1906.- Т. 35, № 3. - C.
1- 44.
. Дорофеюк Н. И., Цэцэгмаа Д.
Конспект флоры водорослей Монголии // Тр. совместной Российско-Монгольской
комплексной экспедиции. Т. 42. М.: Наука, 2002. 283 с.
. Елизарова В.А. (1974)
Содержание фотосинтетических пигментов в единице биомассы фитопланктона
Рыбинского водохранилища. В: Экзерцев В.А. (ред.) Флора, фауна и микроорганизмы
Волги. Рыбинск, ИБВВ АН СССР, с. 46-66.
. Елизарова В.А. (1993)
Хлорофилл как показатель биомассы фитопланктона. В: Пырина И.Л. (ред.)
Методические вопросы изучения первичной продукции планктона внутренних
водоемов. Спб., Гидрометеоиздат, с. 126-131.
. Ермолаев В.И. (1989)
Фитопланктон водоёмов бассейна озера Сартлан. Новосибирск, Наука, 96 с.
. Жизнь растений. В 6-ти т.
Т. 3. Водоросли, лишайники / Под ред. А.А. Федорова. - М.: Просвещение, 1977. -
487 с.
. Загоренко Г.Ф., Кожова О.М.
Растительные обрастания и их роль в продуктивности Братского водохранилища //
Биологическая продуктивность водоемов Сибири. М.: Наука, 1969. - С. 76-79.
. Загоренко Г.Ф., Кожова О.М.
Состав и экологические показатели фитопланктона озера Хубсугул в 1971 г. //
1риродные условия и ресурсы Прихубсугулья (Монгольская Народная Республика).
Иркутск-Улан-Батор, 1973. Вып. 2. С. 329-340.
. Загоренко Г.Ф., Прозоров
В.А. О поясном распределении макрофитов в озере Хубсугул // Природные условия и
ресурсы некоторых районов МНР: Тез. докл. Улан-Батор, 1980. С. 149-151.
Загоренко Г. Ф. Река Селенга // Рыбы Монгольской Народной Республики. М.:
Наука, 1983. С. 45-60.
. Ижболдина Л.А. Макрофиты Южного
Байкала и продукция некоторых видов, доминирующие в литорали района Больших
Котов. «Продуктивность Байкала и антропогенные изменения его природы».
Издательство БГНИИ при ИГУ, 1974.
. Ижболдина Л.А. Макрофиты
Южного Байкала и продукция некоторых видов, доминирующих в литорали района
БольшихКотов //Продуктивность Байкала и антропогенные изменения его природы. -
Иркутск, 1974а.- С.111-116.
. Ижболдина. Л.А. Новые
данные о бентосных водорослях озера Байкал // Новые материалы по фауне и флоре
Байкала, 1976а. -С. 4-13.
. Ижболдина Л.А. Мейо- и
макрофитобентос озера Байкал (водоросли). - Иркутск: изд-во ИГУ. 1990.
. Ижболдина Л.А.
Макрофитобентос открытых побережий озера // Подводные ландшафты Байкала. -
Новосибирск, 1990а. - С. 142-166.
. Ижболдина Л.А. Мейо- и
макрофитобентос озера Байкал (водоросли). - Иркутск, 1990в. - 175с.;
. Ижболдина Л.А. Мейо- и
макрофиты // Флора и фауна водоемов и водотоков Байкальского заповедника
(Аннотированные списки видов). Флора и фауна заповедников. - Вып. 92. - М.,
2001. - с. 15-18.
. Ижболдина Л.А. Атлас и
определитель водорослей фитобентоса и перифритона озера Байкал (мейо- и
макрофиты) с краткими очерками по их экологии. - Новосибирск: Наука, 2007.
. Изместьева Л.Р. (1989)
Связь между биомассой фитопланктона и концентрацией хлорофилла. В: Коновалов
С.М. (ред.) Экология фитопланктона Куйбышевского водохранилища. Л., Наука, с.
199-202.
. Каплина Г.С. Макрозообентос
каменистых грунтов литорали оз. Байкал и его сезонная динамика (данные 1963 -
1968 гг., район Больших Котов)// Продуктивность Байкала и антропогенные
изменения его природы. 1974.
. Кожов М.М. О
видообразовании в озере Байкал // Бюл. МОИП. Отд. Биологии. - М., 1960. - Т.
65. - С. 39-47.
. Кожов М.М. Биология озера
Байкал. - М., 1962. - 315 с.
. Кожова О.М., Загоренко Г.Ф
К вопросу определения первичной продукции … Байкал в 1968 г. // Продуктивность
Байкала и антропогенные изменения его природы; 1974; 153-159.
. Куклин А.П. Макрофитные
водоросли - индикаторы загрязнения речной сети приграничных территорий (на
примере Кыринского района Забайкальского края) // Природоохранное
сотрудничество в трансграничных экологических регионах: Россия - Китай -
Монголия: Сб. науч. мат. - Вып. 2. - Чита: Экспресс-издательство, 2011. - с.
117-121
. Куликова Н.Н., Сайбаталова
Е.В., Козырева Е.И. Химический элементный состав Ulothrix Zonaта (Web. Ет Mohr)
Kutz. залива Большие Коты оз. Байкал. // Материалы II международной конференции
'Биоразнообразие, проблемы экологии горного Алтая и сопредельных регионов:
настоящее, прошлое, будущее'; РИО ГОУВПО 'Горно-Алтайский государственный
университет' : .2010; ():108-112.
. Курейшевич А.В., Сиренко
JI.A., Медведь В.А. Многолетняя динамика содержания хлорофилла а и особенности
развития фитопланктона в Днепродзержинском водохранилище // Гидробиол. журн. -
1999. -35, №3.-С. 49-62.
. Ляшенко О.А. (1991)
Фитопланктон озера Неро. В: Ривьер И. К. (ред.) Современное состояние
экосистемы озера Неро. Рыбинск, ИБВВ АН СССР, с. 10-32.
. Ляшенко О.А. (2004)
Растительные пигменты как показатели биомассы фитопланктона в мелководном
эвтрофном озере. Проблемы региональной экологии. 5: 6-14
. Малый практикум по низшим
растениям / Н.П. Горбунова, Е.С. Клюшникова, Н.А. Комарницкий и др. - М., 1976.
. Мейер К.И., Рейнгард Л.В. К
флоре водорослей оз. Байкала и Забайкалья // Бюлл. МОИП, 1925. - Т. 33. - Вып.
3-4. - С. 201 - 243.
. Мейер К.И. Введение во
флору водорослей озера Байкала // Бюл. МОИП. Отд. Биологии. - М., 1930. - Т.
39., вып.3-4. - С. 179-396.
. Растительные пигменты в
воде волжских водохранилищ/Н.М. Минеева. Отв. Ред. В.Т. Комов.-М.: Наука,
2004.- 156 с.: ил.- ISBN
5-02-033277-1 (в пер.)
. Минеева Н.М. (2004)
Растительные пигменты в воде Волжских водохранилищ. М., Наука, 156 с.
. Минеева Н.М. (2006)
Содержание фотосинтетических пигментов в водохранилищах Верхней Волги
(1994-2003 гг.). Биология внутренних вод. 1: 31-40.
. Минеева Н. М., Щур Л. А.,
Бондаренко Н. А. Функционирование пресноводного фитопланктона при разной
обеспеченности энергетическими ресурсами // Тез. докл. IV Междунар. науч. конф.
«Современные проблемы гидроэкологии». - Россия, Ст.-Петербург. - 2010. - С.
120.
. Окунева Г.Л. Сезонные
изменения мезобентоса на каменистой литорали (район пос. Большие Коты) //
Продуктивность Байкала и антропогенные изменения его природы. 1974.
. Окунева Г.Л. Мезо- и микробентос
в районе Больших Котов (Южный Байкал) // Новые материалы по фауне и флоре
Байкала. - Иркутск, 1976.
. Пырина И.Л. (1992)
Содержание хлорофилла и продуктивность фитопланктона озера Плещеево. В:
Семерной В.П. (ред.) Факторы и процессы эвтрофикации озера Плещеево. Ярославль,
ЯрГУ, с. 18-28.
. Рабинович Е. Фотосинтез.
Пер. с англ. М.: Изд-во иностр. лит., 1951. Т. 1. 648 с.
. Россолимо Л.Л.
Температурный режим озера Байкал // Тр. Байкал. лимн. станции.-1957.-С.551.
. Сигарева Л.Е., Ляшенко О.А.
(1991) Пигментные характеристики фитопланктона озера Неро. В: Ривьер И. К.
(ред.) Современное состояние экосистемы озера Неро. Рыбинск, ИБВВ АН СССР, с.
32-53.
. Сигарева Л.Е., Ляшенко О.А.
(2004) Значимость пигментных характеристик фитопланктона при оценке качества
воды. Водные ресурсы. 4: 475-480Сигарева и Ляшенко, 2004;
. Скабичевский А.П. о
распределении донной растительности в губах Аяя, Фролиха и Лаканда на Байкале
// Изв. Биол. - геогр. НИИ при иркут. Ун-те. - 1934. - Т. 4,вып. 1. - С. 103 -
115.
. Скабичевский А.П. Заметки
по альгологии Байкала //Изв. Биол. - геогр. НИИ при иркут. Ун-те. - 1936. - Т.
7,вып. 1-2. - С. 32 - 41.
. Скабичевский А.П. О
распределении донной растительности Байкала в окрестностях Больших Котов //
Бюл. МОИП. Отд. Биол. - 1966. - Т. 71, № 6. - С. 108-119.
. Тарчевский И.А. Основы
фотосинтеза/И.А. Тарчевский. - 1977.
. Трифонова И.С. (1979)
Состав и продуктивность фитопланктона разнотипных озёр Карельского перешейка.
Л., Наука, 168 с.
. Трифонова И.С., Десортова
Б. (1983) Хлорофилл как мера биомассы фитопланктона в водоёмах разного типа.
Гидробиологические процессы в водоемах. Л., Наука, с. 58-80.
. Туровцев В.Д., Краснов В.С.
Т 88 Биоиндикация: Уеб. Пособие. - Тверь: Твер. гос. ун-т, 2004. - 260 с.).
. Унифицированные методы
изучения вод. 1977. Индикаторы сапробности, 3. Изд-во СЭВ: 42 c.
. Унифицированные методы
исследования качества вод, ч. 3. Методы химического анализа вод. Методы
биологического анализа вод 1983г.//Совещание руководителей водохозяйственных
органов стран-членов СЭВ. 1990. - 371 с.
. Формирование растительного
перифитона Братского водохранилища : / Загоренко Г.Ф., Г.Ф.Загоренко; Иркутский
гос.ун-т .-Иркутск : ***, 1970 .- 23c . ББК 28.082.
. Шимараев М.Н. Элементы
теплового режима озера Байкал. - Новосибирск, 1977.- 147 с.
. Щадрина Л.А.
Экотоксикологическое картирование морских побережных вод методом
биотестирования //Гидробиол.журн. 1997. Т. 33, №6. С50-55.
. Эдвардс Дж., Уокер Д.
Фотосинтез С3- и С4-растений: механизмы и регуляция. М. Мир, 1986.
72. Ahlgren G. Lake
Norrviken, a eutrophicated Swedish Lake // Phytoplankton and its production.-
Schweiz. L. fur Hydrologie, 1970.-Bd. 32.-H.2.-S. 353-395.
. Antrium L.D., Thom R.
M., Gardiner W.W. et al. Effects of petroleum products on bull help Nereocystis
luetreana // Mar. Biol. 1995. Vol.
122, N1. P.
32-31. Asha Udayamali Meegolle Lokuhewage, 2011.
74. Benhissoune, S.,
Boudouresque, C.-F. & Verlaque, M. (2001). A
checklist of marine seaweeds of the Mediterranean and Atlantic coasts of
Morocco I. Chlorophyceae Wille s. l.. Botanica Marina 44:
171-182.
. Biggs, J.F. &
Close, M.E. 1989. Periphyton biomass dynamics in gravel bed rivers: the
relative effects of flows and nutrients. Freshwater Biology 22: 209-231;
76. Boney, A.D. (1981)
Mucialge: the ubiquitous algal attribute. Br.phycol. J. 16: 115-132.
. Bold, H.C. and M.J.
Wynne, 1985. Introduction to the Algae (Structure and Reproduction). 2nd
Edn., Prentice-Hall Inc., Englewood Cliffs, USA.
. Canfield D.E.Jr.,
Linda SB., Hodgson L.M. (1985) Chlorophyll-biomass-nutrient relationships for
natural assemblages of Florida phytoplankton. J. AWRA. 3: 381-391.
79. Сommunities
in the ultrapure ice of Lake Baikal: distinctive characteristics and adaptive
strategies of the inhabitants/ Osipova S. [et al]. // Proc. 20th IAHR Intern.
Symp. on Ice, 14-17 June 2010, Lahti, Finland (ISBN 978-952-10-5979-7), paper №
40;
. Croasdale, H.T.
(1973). Freshwater algae of Ellesmere Island, N.W.T. (exclusive of diatoms and
flagellates). National Museum of Canada Publications in Botany 3: 1-131,
28 pls, 3 graphs.
81. Curry, M.G., Everitt,
B. & Widrine, M.F. 1981. Haptobenthos on shells of living freshwater clams
in Louisiana. Journal of Biology 39: 56-63; Power, M.E. 1990a. Effects
of fish in river food webs. Science 250:811-814; Power, M.E. 1992. Hydrologic
and trophic controls of seasonal algal blooms in northern California rivers. Archivs
fur Hydrobiologie 125: 385-410.
82. Day, S.A., Wickham,
R.P., Entwisle, T.J. & Tyler, P.A. (1995). Bibliographic check-list of
non-marine algae in Australia. Flora of Australia Supplementary Series
4: vii + 276.
. Desortova B. (1981)
Relationship between chlorophyll-a concentration and phytoplankton biomass in
several reservoirs in Czechoslovakia. Int. Revue ges. Hydrobiol. 2: 153-169.
. Dodds, W.K. 1991a.
Micro-environmental characteristics of filamentous algal communities in flowing
freshwaters. Freshwater Biology 25: 199-209.
85. Dodel : Jahrbnch Wiss.
Bot., x, 1876.
. Dudley, T.L. &
D’Antonio, C.M. 1991. The effects of substrate texture, grazing, and
disturbance on macroalgal establishment in streams. Ecology 72(1): 297-309.
87. Evans, D. &
Stockner, J. G. 1972. Attached algae on artificial and natural substrates in
Lake Wennipeg, Manitoba. J. Fish, Res. Board Can. 29:31-44;
88. Canfield D.E.Jr., Linda
SB., Hodgson L.M. (1985) Chlorophyll-biomass-nutrient relationships for natural
assemblages of Florida phytoplankton. J. AWRA. 3: 381-391.
. Felip M., Catalan J.
(2000) The relationship between phytoplankton biovolume and chlorophyll in a deep
oligotrophic lake: decoupling in their spatial and temporal maxima. J. Plankton
Res. 1: 91-106.
90. Fisher, S.G., Gray,
L.J., Grimm, N.B. & Busch, D.F. 1982. Temporal succession in a desert
stream ecosystem following a flash flooding. Ecological Monographs 52:
93-110.
91. Foy R.H. (1987) A
comparison of chlorophyll-a and carotenoid concentrations as indicators of
algal volume. Freshwater Biology. 17: 237-250.
92. Gallardo, T., Gómez
Garreta, A., Ribera, M.A., Cormaci, M.,
Furnari, G., Giaccone, G. & Boudouresque, C.-F. (1993). Check-list of
Mediterranean Seaweeds, II. Chlorophyceae Wille s.l.. Botanica Marina
36: 399-421, 1 fig, 1 table.
. Geitler, L., 1977: Das
rhizoid von Ulothrix zonata (Chlorophyceae). - Pl. Syst. Evol. 128, 71-78.
94. Graham, J.M.,
Kranzfelder, J.A. and M.T. Auer. 1985. Light
and temperature as factors regulating seasonal growth and distribution of Ulothrix
zonata (Ulvophyceae). J. Phycol. 21: 228-234.
. Graham, J.M., Graham,
L.E. and J.A. Kranzfelder. 1985. Light, temperature and photoperiod as factors
controlling reproduction in Ulothrix zonata (Ulvophyceae). J.
Phycol. 21: 235-239.;
. Granberg K., Harjula
H. (1982) On the relation of chlorophyll-a to phytoplankton biomass in some
Finnish freshwater lakes. Arch. Hydrobiol. Beih. 16: 63-75.
98. Gross, I. :
Entwicklungsgeschichte, Phasenwecbsel nnd Sexnalitat von Ulothrix, Archiv f.
Protistenk., Bd. 72, Heft a.
. Guiry, M.D. &
Guiry, G.M. 2012. AlgaeBase. World-wide electronic publication, National
University of Ireland, Galway. http://www.algaebase.org; searched on 16 June
2012.
. Hansen, G.I. (1997). A
revised checklist and preliminary assessment of the macrobenthic marine algae
and seagrasses of Oregeon. In: Conservation and Management of Native Flora
and Fungi. (Kaye, T.N., Liston, A., Love, R.M., Luoma, D.L., Meinke, R.J.
& Wilson, M.V. Eds), pp. 175-200. Corvallis: Native Plant Society of Oregon.
. Hawes I, Schwarz A-M
(1996) Epiphytes from a deep-water characean meadow in an oligotrophic New
Zealand lake: species composition, biomass and photosynthesis. Freshwater Biol
36:297-313.
. Herbst, R.P. 1969.
Ecological factors and the distribution of Cladophora glomerata in the Great
Lakes. Am. Mid. Nat. 82:90-98.;
103. Hill WR (1996) Effects
of light. In: Stevenson J, Bothwell ML, Lowe RL (eds) Algal ecology -
freshwater benthic ecosystems. Academic, San Diego, pp
121-148.
. Hillebrand H.,
Durselen C-D., Kirschtel D., Pollingher U., Zohary T. (1999) Biovolum
calculation for pelagic and benthic microalgae. J. Phycol. 35: 403-424.
. Hooten R.L. Carr R.S.
Development and application of a marine sediment pose- toxity test using Ulva
fasciata zoospores // Environ. Toxicol. And Chem. 1998. Vol. 17, N 5. P.
932-940.
. Hu, H. & Wei, Y.
(2006). The freshwater algae of China. Systematics, taxonomy and
ecology. pp. [4 pls of 16 figs], [i-iv], i-xv, 1-1023.
107. Javornicky P. (1974) The
relationship between productivity and biomass of phytoplankton in some
oligotrophic waterbodies in the German Democratic Republic. Limnologica 9:
181-195.
. John, D.M. (2002). Orders
Chaetophorales, Klebshormidiales, Microsporales, Ulotrichales. In: The
Freshwater Algal Flora of the British Isles. An identification guide to
freshwater and terrestrial algae. (John, D.M., Whitton, B.A. & Brook,
A.J. Eds), pp. 433-468. Cambridge: Cambridge University Press.
109. Jonsson GS (1987) The
depth-distribution and biomass of epilithic periphyton in Lake Thingvallavatn.
Arch Hydrobiol 108:531- 547.
. Jonsson GS (1992)
Photosynthesis and production of epilithic algal communities in Thingvallavatn.
Oikos 64:222-240.
. Kairesalo, T. 1980.
Diurnal fluctuations within a littoral plankton community in oligotrophic Lake
Paajarvi, southern Finland. Freshwater Biology, 10;
533-537.
. Kalchev R.K., Beshkova
M.B., Boumbarova C.S., Tsvetkova R.L., Sais D. (1996) Some allometric and
non-allometric relationship between chlorophyll-a and abundance variables of
phytoplankton. Hydrobiologia. 341: 235-245.
. Kentaro Nozaki,
Hiroshi Morino. Hiroyuki Munehara,Valentina G. Sideleva · Katsuki Nakai,Mitsugu
Yamauchi, Olga M. Kozhova, Masami Nakanishi (2002) Composition, biomass, and
photosynthetic activity of the benthic algal сommunities
in a littoral zone of Lake Baikal in summer. / Limnology 3:175-180.
. Kingston J.C., Lowe
R.L., Stoermer E.F., Ludewski T. (1983) Spatial and temporal distribution of
benthic diatoms in northern Lake Michigan. Ecology 64:1566-1580.
115. Kützing, F.T. (1843). Phycologia
generalis oder Anatomie, Physiologie und
Systemkunde der Tange... Mit 80 farbig gedruckten Tafeln, gezeichnet und gravirt
vom Verfasser. pp. [part
1]: [i]-xxxii,
[1]-142, [part 2:]
143-458, 1, err.], pls
1-80.
. Lee E.R. 1999.
Phycology. Cambridge University Press, Cambridge, UK. 614 pp.
. Lewis, L. A.
& McCourt, R. M. (2004). "Green algae and the origin
of land plants". American Journal of Botany 91
(10): 1535-1556.
. Lind E.M. 1932. A
contribution to the life cycle history and cytology of two species of Ulothrix.
Ann. Bot. 46: 711-725.
. Loeb ST, Reuter JE
(1981) The epilithic periphyton community: a fivelake comparative study of
community productivity, nitrogen metabolism and depth-distribution of standing
crop. Verh Internat Verein Limnol 21:346-352.
. Lokhorst G.M. &
Vroman, M. 1972. Taxonomic study of three freshwater Ulothrix species. Acta.
Bot. Neerl. 21:449-80.
121. Lokhorst G.M. &
Vroman, M., 1974: Taxonomic studies on the genus Ulothrix (Ulothrichales,
Clorophyceae) III.- Acta Bot. Neerl.23, 561-602.
. Lorenzen G.J. (1967)
Determination of chlorophyll and pheopigments: spectrophotometric equations.
Limnology and Oceanography. 2: 343-346.
. Mattox, K.R. &
Stewart, K.D. 1984. Classification of the green algae: a concept based on
comparative cytology. Pp. 29‐72,
In Irvine, D.E.G. & John, D.M. (eds.), The systematics of Green
Algae. The Systematics Association, Special Vol. 27, Academic Press, London.
. Moore, J.W. 1977. Some
factors effecting algal densities in a eutrophic farmland stream.Oecologia 29:
257-267.
. Moore, L. F. 1978.
Attached algae at thermal generating stations- the effects of temperature on
Cladophora. Int. Verein.
Limnol., Verh.
20: 1727- 33.
126. Nicholls K.H., Dillon
P.J. (1978) An evaluation of phosphorus-chlorophyll-phytoplankton relationships
for lakes. Int. Revue ges. Hydrobiol. 2: 141-154.
127. Nielsen, R.,
Kristiansen, A., Mathiesen, L. & Mathiesen, H. (1995). Distributional index
of the benthic marine macroalgae of the Baltic Sea area. Acta Botanica
Fennica 155: 1-70.
128. Owens. T.G., Falkowski
P.G. and Whitledge T.E., 1980. Dicl periodicity in cellular chlorophyll
contentin marine diatoms. Mar. Biol. 59:71-77.
. Parker, R.D.R. 1979.
Distribution of filamentous green algae in near- shore periphyton communities
along the north shore of Lake Superior. Am. Mid. Nat. 101:326-32.
130. Parker, R.D.R. &
Drown, D.B. 1982. Effects of phosphorus enrichment and wave simulation on
populations of Ulothrix zonata from northern Lake Superior. J.
Great Lakes.
Res. 8:16-26.
131. Pascher :
Susswasserflora. Chlorophyceae, 3.
. Riemann B., Simonsen
P., Stensgaard L. (1989) The carbon and chlorophyll content of phytoplankton
from various nutrient regimes. J. Plankton Res. 5: 1037-1045.
133. Rippka, R. (1988).
Recognition and identification of cyanobacteria. Methods Enzymol.
. Pfeifer, R.F. &
McDiffett, W.F. 1975. Some factors affecting primary productivity of
streamriffle communities. Hydrobiologia 75: 306-317.
135. Prescott, G.W. (1961). Algae
of the western Great Lakes area with an illustrated key to the genera of
desmids and freshwater diatoms. pp. xiii + 977. Dubuque, Iowa: Wm. C.
Brown.
. Power, M.E. &
Stewart, A.J. 1987. Disturbance and recovery of an algal assemblage following
flooding in an Oklahoma stream. American Midland Naturalist 117: 333-345.
137. Sarma, Y.S.R.K., 1962:
Contributions to the karyology of the genus Ulotrichales - I. Ulothrix. -
Phycologia 2, 173-183.
. Sluiman, H.J. (1989).
The green algal class Ulvophyceae. An ultrastructural survey and
classification. Cryptogamic Botany, 1: 83-94.
. Sluiman, H.J.,
Stewart, K.D. & Mattox, K.R. (1980). The ultrastructure of the scaly
zoospore of Ulothrix zonata: its taxonomic significance. Journal of
Phycology: 40.
140. Smith, R.F. &
Ludwig, H.F. 1968. Eutrophication mechanisms at Lake Tahoe II. Source of an
endogenous Ulothrix bloom on the south shore, July, 1967. Water Res. 2:625- 30.
141. Shyam, R., Saxena, P.
N., Morphological and cytological investigations of Ulothrix zonata and
taxonomy of the related species in: Plant systematics and evolution,
ISSN 1615-6110, Vol. 135 (3/4. 1980), p. 151-158.).
142. Shussning, B., 1930: Die
mitotische Kernteilung bei Ulothrix zonata Kützing. Z.
Zellforsch. 10, 642-650.
. Stevenson RJ, Stoermer
EF (1981) Quantitative differences between benthic algal communities along a
depth gradient in Lake Michigan. J Phycol 17:29-36.
. Tadros M. G., Phillips
J., Patel H., Pandiripally V. Differential response of marine diatoms to
solvents // Bull. Environ. Contam.and Toxicol. 1995. Vol. 54, N 6. P. 924-929.
145. Taskin, E.,
Öztürk, M., Kurt, O. & Öztürk, M. (2008). The
check-list of the marine algae of Turkey.
pp. [i-ii]-[1]-87. Manisa, Turkey: Ecem Kirtasiye.;
. Temporal variation in
fatty acid composition of Ulotrix zonata (Chlorophyta) from ice benthic
communities of Lake Baikal/ Osipova S. [et al]. // Phycologia, 2009. V. 48(2),
P.130-135.
. Tolstoy A. 1979)
Chlorophyll a in relation to phytoplankton volume in some Swedish lakes. Arch.
Hydrobiol. 2: 133-151.
. Voros L., Padisak J.
(1991) Phytoplankton biomass and chlorophyll-a in some shallow lakes in central
Europe. Hydrobiologia. 215: 111-119.
149. Weber F., Mohr, D.M.H.,
1804: Naturhistorische Reise durch einen Theil Schwedens.- Göttingen.);
150. Zarina, A.,
Masud-ul-Hasan and M. Shameel. 2005. Taxonomic study of Vaucheriophyta Shameel
from certain areas of the Punjab and NWFP, Pakistan. Int. J. Phycol.
Phycochem., 1: 159-166.
. Zarina, H.R.,
Masus-ul-Hasan & Shameel, M. (2007). Taxonomic study of Chlorophyta from
Lahore, Pakistan. International Journal of Phycology and Phycochemistry
3(2): 173-182.