Бабочки дневные
ДИПЛОМНА РОБОТА
Еколого-фаунстичний аналіз грунтової мезофауни Рожищенського
району
ЗМІСТ
ВСТУП
РОЗДІЛ 1.
Теоретичні засади дослідження
.1 Огляд літератури
.2 Опис району вивчення
РОЗДІЛ 2. Матеріал та методика
дослідження
РОЗДІЛ 3. Аналіз результатів дослідження
.1 Еколого-фауністична характеристика
грунтової мезофауни
Рожищенського району
.2 Хорологія грунтової мезофауни у районі дослідження
.2.1 Характеристика типових лісів
Рожищенського району
.2.2 Поширення грунтової мезофауни у
соснових лісах
.2.3 Поширення грунтової мезофауни у
березових лісах
ВИСНОВКИ
СПИСОК ВИКОРИСТАНОЇ ЛІТЕРАТУРИ
ВСТУП
екологічний фауністичний мезофауна грунтовий ліс
Ґрунт - природне біокосне тіло, поряд з такими
його складовими, як тверда, рідка і газоподібна фази, містить і живу частину.
До складу ґрунтової біоти входять представники всіх царств живої природи.
Ґрунтове населення не тільки живе в ґрунті,
використовуючи його як середовище існування, але й активно трансформує його.
Тому цілком закономірне твердження, що ґрунт - це середовище існування й
одночасно продукт діяльності живих істот, що його населяють.
Ґрунтова біота - найбільш динамічна ланка
екосистеми, яка зазнає періодичних змін і, у той же час, що володіє певним
гомеостазом завдяки наявності механізмів регуляції і контролю.
Грунтові біоценози представлені
найрізноманітнішими безхребетними тваринами, а саме: найпростішими
(саркодовими, джгутиковими, інфузоріями), нематодами, коловертками, олігохетами
(люмбрицідами і енхитреїдами), черевоногими молюсками, хеліцеровими (павуками,
кліщами, косариками, псевдоскорпіонами), ракоподібними, багатоніжками,
комахами. При урахуванні особливостей життя і впливу на грунт тварин різних
розмірів їх поділяють на три групи: мікрофауну, мезофауну та мегафауну.
Мезофауна є важливим компонентом біогеоценозів.
Вона займає ведуче місце в зоомасі в складі ценозу, істотно впливає на процеси
трансформації органічних решток, міграції хімічних елементів в горизонтах
ґрунтового профілю, ґрунтоутворення, стимулювання біологічних процесів та
сукцесії організмів в біогеоценозах ґрунту. Без обліку мезофауни населення
ґрунту неможливо скласти загальний кадастр тваринного світу. Разом з тим,
мезофауна ґрунту включає значну кількість лісових і сільськогосподарських
шкідників, а тому має велике практичне значення.
Серед представників мезофауни ґрунтових
безхребетних тварин є багато стенобіонтних видів. Власне вони відповідають
вимогам індикації стану оточуючого середовища і можуть служити об‘єктами
моніторингу.
Розв‘язати завдання в напрямку біоіндикаційних
цілей можна лише на основі дослідження угрупування ґрунтових безхребетних в
цілому, в тому числі, представників мезофауни.
Актуальність дослідження зумовлена недостатнім
рівнем відомостей про фауністичний склад, особливості екології та поширення
ґрунтової мезофауни Рожищенського району.
Мета роботи: вивчити еколого-фауністичні
особливості ґрунтової мезофауни району дослідження.
В роботі ставились такі завдання:
. Проаналізувати літературні джерела та
ознайомитись з історією вивчення ґрунтової мезофауни.
. Розглянути фізико-географічні умови
Рожищенського району
. Встановити фауністичний склад ґрунтової
мезофауни у районі дослідження.
. Вивчити екологію ґрунтової мезофауни у
Рожищенському районі.
. Виявити особливості поширення мезофауни у районі
вивчення.
Об’єктом дослідження є ґрунтова мезофауна
Рожищенського району. Предметом дослідження є фауна, екологія, особливості
поширення ґрунтової мезофауни у районі дослідження.
Наукова новизна дослідження. Вперше вивчено
видовий склад, особливості екології та хорології грунтової мезофауни у
Рожищенському районі.
Практичне значення дипломної роботи полягає в
тому, що його результати можуть бути використанні у професійній діяльності
педобіологів, ґрунтознавців, агрономів, працівників сільського господарства та
лісництва, а також у навчальному процесі. Дані роботи можна використовувати
також при проведенні інтродукції та акліматизації окремих видів у відповідності
до типів ґрунтів.
Структура роботи. Дипломна робота складається із
вступу, трьох розділів, висновків, списку використаної літератури, який містить
57 джерел. Роботу представлено на 72 сторінках друкованого тексту.
РОЗДІЛ 1. ТЕОРЕТИЧНІ ЗАСАДИ ДОСЛІДЖЕННЯ
.1 Огляд
літератури
З другої половини ХІХ століття і до наших днів зі
сторони вітчизняних і зарубіжних зоологів та ґрунтознавців значно зріс інтерес
до життєдіяльності ґрунтових тварин.
Важливою ланкою грунтової мезофауни є олігохети.
Зацікавленість природодослідників ґрунтовими олігохетами родини люмбріцід була
започаткована ще в першій половині ХІХ століття. Про це свідчать повідомлення у
періодичних виданнях Галичини (Західна Україна) “Ziemianin” та “Prirodnik
Lwowski” [49].
Перші відомості про люмбріцід України, які мали
науковий характер, належать М.М. Кулагіну. Зазначимо, що на даний час систематика
люмбріцід була слабо розроблена. На другу половину ХІХ століття припадає
найбільша кількість описів нових видів.
М.М. Кулагін при описах дощових червів
користувався лише зовнішніми морфологічними ознаками, а тому зараз важко
встановити, з якими саме видами він мав справу. В працях він вказує вперше для
України Lumbricus rubellus, Allolobophora foetida (=Eisenia foetida), A. cjgnea
(= A. rosea), A. carnea (A. caliginosa) [18].
На подальший розвиток досліджень ґрунтових
олігохет вплинули роботи Я. Нузбаума. Ці публікації на тлі попередніх робіт
відрізнялись новими підходами у вивченні люмбріцід. Вперше автор подає зведення
видового складу дощових червів Західної України. Фауністичний склад люмбріцід
за його описами представлений 14 видами, 7 родами. Це були перші зведення
видового складу люмбріцід західних областей України та східної Польщі [53].
А. Яворовський при дослідженні тваринного
населення криниць виявив п’ять видів олігохет, серед них Lumbricus terrestris
[56].
Значний доробок у дослідженні фауністичного
складу ґрунтових олігохет Європи і Азії належить В. Міхаельсену. Ним була
розроблена і запропонована систематика олігохет, яка привернула до себе значну
увагу багатьох систематиків. В монографічній роботі В. Міхаельсен наводить
шість видів дощових червів, що доповнили фауністичний склад люмбріцід України
[50].
В першій половині ХХ століття дослідження
ґрунтових олігохет мали фауністичне спрямування. Необхідно відзначити роботи Л.
Андрусова, Cognetti de Marti, L. Cernosvitov, які значно розширили відомості
про поширення люмбріцід у різних фізико-географічних областях України [2].
В 20-30 р.р. ХХ ст. оригінальні дослідження
олігохет Польщі, західної України і Білорусії проводив А. Мошинський. Можна
вважати, що перша робота, яка була присвячена олігохетам Західного Полісся,
належить Мошинському. Вона мала комплексний характер. В ній вказувалося на
знаходження люмбріцід [49].
Оригінальними виявились роботи Й.Й. Малевича,
А.Ф. Кіпенварлиць. Вони стосувалися південної частини Білорусії, територія якої
є суміжною з Житомирським Поліссям. Як покажуть подальші дослідження
фауністичного складу дощових червів, Й.Й. Малевич вже у 1937 р. найбільш повно
встановив фауністичний склад люмбріцід біоценозів південної частини Білорусії
[20].
По закінченню другої світової війни отримали
розвиток еко-фауністичні дослідження дощових червів. В 50-60 рр. ХХ століття в
Україні плідно розвивав цей напрямок А.І. Зражевський. Його праця була
спрямована на вивчення видового складу дощових червів лісових масивів України,
їх приуроченість до певного рослинного покриву, а також показано їх вплив на
ріст рослин та ґрунтоутворювальну роль у конкретних біоценозах. Автор також
розглядав окремі питання живлення і поширення даних грунтових тварин в
залежності від абіотичних факторів. А.І. Зражевським була розроблена методика
дослідження діяльності дощових червів під наметом лісу. [14].
Найбільш повний видовий склад ґрунтових олігохет
родини Lumbricidae західного регіону України представлений в роботах В.В.
Іванціва та Л.В. Бусленок, які вивчали структурно-функціональну організацію
комплексів грунтових олігохет [5; 13].
Необхідно зазначити, що в повоєнний час
спеціальні дослідження дощових червів Житомирського Полісся були розпочаті лише
з 80 років ХХ століття. В роботі Е.В. Шапошникової наводяться скупі відомості
про фауністичний склад люмбріцід України, в тому числі, Житомирського Полісся
[40].
Енхитреїди, як таксономічна група, до ХХ століття
практично не вивчалась. Наявні скупі відомості в роботах польських зоологів
кінця XIX - початку XX ст. про поширення окремих видів в межах України.
Потрібно відзначити, що дослідження мали фауністичний характер і являлись
початковим етапом пізнання групи. Лише з 80-х років набувають розвитку
екологічні дослідження видів, їх трофічних зв`язків та значення в ґрунтових
ценозах.
Дослідження стосовно ґрунтової мезофауни України
здійснювали такі науковці як: О.М. Піндрус, П.П. Яворівський, О.В. Лукаш [24].
Б.Р. Стриганова в роботі “Комплексы почвообитающих
беспозвоночных в пойме среднего течения Днестра” вказує на наявність у регіоні
дослідження основних комплексів грунтових тварин, аналізує особливості
поширення та екологію даних організмів [33].
При вивченні безхребетних тварин заплавних
біотопів басейну ріки Тетерев В.Г. Надворний відзначає особливості поширення
ґрунтових організмів, в тому числі, ґрунтової мезофауни по профілю заплави.
Кількісна характеристика участі ґрунтової
мезофауни в розкладі рослинних решток наведена в роботах Й.З. Царика, Е.В.
Шапошникової, Т.А. Казидуба, А.К. Ткаченка, І.Л. Гавриша, І.С Ейтминавичюте
[39; 40; 41].
Грунтових безхребетних, зокрема, мезофауну
лісових біогеоценозів досліджували М.С. Гіляров, Ю.І. Чернов та ін. В цих
роботах дається характеристика зонального поширення тварин ґрунтів. При цьому
приводяться параметри їх чисельності, зоомаси і метаболізму [8; 9; 10; 36].
Слід відзначити цілий ряд робіт присвячених
впливу на мезофауну інтенсивного забруднення ґрунтів промисловими викидами,
зокрема, це відображено у монографіях О.М. Кунах, О.Є. Пахомова, Е.І. Хотько та
ін [27; 38].
Найперші відомості з фауни молюсків родин
Melanopsidae, Lithoglyphidae, Bithyniidae Правобережної України датуються
першою половиною ХІХ століття. Зокрема, вивченням даних груп грунтової
мезофауни займалися такі вчені як І.В. Ейхвальд та Л.В. Сімешко. Тут у списках
молюсків для Литви, Полісся та Поділля знаходимо 5 видів з вищезазначених родин
- Fagotia acicularis (Fèr.), F. esperi (Fèr.), Lithoglyphus naticoides (C. Pfeif.), L. fuscus (C.
Pfeif.), Bithynia tentaculata (Linné.) [46].
У другій половині ХІХ ст. відомості про згаданих
молюсків стали з’являтися частіше. Списки прісноводних молюсків Буковини та
Галичини опублікував О. Завадський, вказуючи також п’ять видів вищезгаданих молюсків.
Чимало відомостей про фауністичний склад,
особливості екології грунтової мезофауни наявні в працях інших дослідників
цього періоду, а саме G. Belke, Е. Eichwald, J. Bąkowski, С. Westerlund [44; 45; 46; 57]. Істотне доповнення до
списку видів ми знайшли в монографії F. Baker, в якій зазначено Bithynia
troscheli, що раніше ніким не згадувався [43].
Найсуттєвішим внеском у вивчення екології та
поширення Melanopsidae, Lithoglyphidae та Bithyniidae стали в цей час роботи Й.
Бонковського, публікації якого мають узагальнюючий характер стосовно
досліджуваної групи тварин [44].
Особливо інтенсивно розвиваються ентомологічні
дослідження у другій половині ХІХ ст. Об’єктами ентомологічних досліджень
стають ті види, які приносять безпосередню або опосередковану шкоду людям. В
результаті чого було проведено чимало робіт по вивченню комах, що пошкоджують
лісові, городні, польові рослини. Все це призвело до широкого поширення
прикладних напрямків ентомології, а отже і поглиблення відомостей стосовно
грунтової мезофауни.
Вперше список видів родини златок зустрічається в
каталозі жуків околиць м. Самбір, у якому вказано 21 вид з даними про місця
збору. Пізніше М. Новицький для фауни Галичини вказав 34 види златок. М.
Ломницький (М. Lomnicki) у каталозі жуків Галичини вказав 49 видів златок.
Дослідником було проведено каталогізацію комах з колекцій музею В. Дідушицьких,
складено видові списки жуків Львова, видано каталог твердокрилих Польщі, куди
були включені всі знайдені ним види [48].
Ж. Круль для Розточчя вказав 10 видів златок.
Окрім того, дослідником була зроблена спроба зоогеографічного аналізу: виділені
північноєвропейські, гірські і подільські елементи. М. Рибінський опублікував
фауністичний список жуків, зібраних на території Поділля, який включає 26 видів
златок із детальним описом місцевостей збору [19].
В. П'ятакова доповнила видовий список жуків
Східного Поділля, складений Л. Хільдтом, і вказала для регіону 15 видів златок.
Ш. Тененбаум вперше для фауни Полісся вказав Agrilus aurichalceus Redt., A.
macroderus Ab. і виявив нові місцезнаходження в регіоні для Anthaxia deaurata
(Gm.) і Agrilus graminis. Р. Кунтце та Й. Носкевич для бупрестідофауни вказали
12 видів. Дослідниками наведено кормові рослини личинок, характерні для імаго
біотопи й поширення видів у Палеарктиці. Для всіх місцевостей збору подано
детальний опис [19].
В. Лазорко для 7 видів златок вказав нові місця
знахідок на території України, а також зробив спробу внести ясність у
синоніміку окремих таксонів.
Чеські вчені проводили дослідження стосовно
впливу сірчаного ангідриду на комах-фітофагів. Таким чином було встановлено, що
соснові ліси піддані дії даних викидів, швидко деградують, що приводить до їх
вимушеної вирубки і штучному насадженню дрібнолистях лісів. В той же час в
Польщі С. Віацковський, показав, що грунтова мезофауна в зоні промислового
забрудення пошкоджує значно більше коренів рослин, порівняно із шкідливістю її
на контрольних площах.
В Білорусії вчені займалися дослідженням впливу
меліорації на ґрунтових безхребетних. Зокрема, це: І.Т. Арзамасов, В.М,
Веремеєв М.С. Долбик, Е.І. Хотько, Т.М. Шевцова. Було встановлено основні
закономірності сукцесій грунтової фауну, які відбулися в результаті
антропогенного впливу [4; 6; 38].
Впливом промислових викидів на ґрунтових
безхребетних займались такі дослідники як Е. Хотько, С. Ветрова. В даних
роботах узагальнено результати вивчення змін, що проходять в структурі
безхребетних, які існують в соснових лісах, під впливом забруднень промисловими
викидами та зроблений прогрноз змін на найближчий час [38].
В Україні, зокрема, в західному регіоні вивчався
вплив антропогенних навантажень на соснові масиви Карпат. Однак ґрунтова фауна
біогеоценозів окремих заповідних територій України, зокрема, Рожищенського
району вивчена недостатньо. Що послужило причиною дослідження грунтової
мезофауни Черемського природного заповідника.
.2 Опис
району вивчення
Територія Рожищенського району розташована
в межах Подільської окраїни Східно-Європейської платформи, фундамент якої
складений інтенсивно дислокованими кристалічними породами протерозою,
розбитими системою великих розломів на окремі блоки, що опущені або підняті на
неоднакову висоту.
Рожищенський район
розташований на границі степової (Волино-Подільське плато) і поліської
(Волинська низовина) частини Волині. Волинське Полісся складає меншу частину
значної області Полісся і являє собою одноманітну низовину з окремими
підвищеннями, в деякій мірі заболочену.
Волинське плато і
Волинське Полісся відрізняється одне від одного в геологічному відношенні тим,
що перше покрите лесом, друге - піщано-суглинистими утвореннями. В даному районі
лесове плато переходить поступово в піщано-суглинисту область Полісся.
Утворення рельєфу Полісся
почалося ще в девонський період. Остаточно він сформувався в льодовиковий і
післяльодовиковий період. Піщані, супіщані і глинисті відклади, залишені при таненні
льодовика, в процесі вивітрювання і перевідкладення талими водами створили
сучасний рельєф даної частини Полісся [7].
При слабовираженому
мікрорельєфі цей район характеризується сильно розвинутим мезорельєфом.
Останній має чітко виражений акумулятивний характер: його нерівності утворені
нерівномірними відкладами льодовикових осадів.
В геологічній будові
району беруть участь палеозойські, мезозойські і кайнозойські відклади.
Палеозойські відклади залягають на значній глибині, над ними знаходяться мезозойські
шари. Із останніх особливо широко розвинуті крейдові відклади. Площа крейдових
відкладів представлена породами сіноманського, туринського і сінонського
ярусів, нижньокрейдові відклади виражені тільки альбськими шарами [25].
Сінонські відклади представлені
пісками в верхній частині з горизонтом кремнію і роговиків. Туронський ярус
виражений піщаною крейдою і мергелями. Сінонські утворення складені мергелями,
крейдою, щільними вапняками і опоками. На докрейдових і крейдових відкладах
залягають четвертинні породи, представлені пісками, супісками, суглинками і
т.д.
Територія району
характеризується слабохвилястим рельєфом і невеликим нахилом на північ.
Мезорельєф представлений невисокими підвищеннями з пологими схилами до 2-3°.
Підвищенні місця мають форму широких невисоких плато. Різниця в коливаннях між;
підвищеннями і пониженнями не перевищує 6-10 м. на понижених ділянках території
дуже поширений мікрорельєф. Тут різниця між підвищеннями і пониженнями в межах
1 м. дуже розвинутий мікрорельєф в північній частині району, у формі блюдець,
які в більшості заболочені. Тому заболочені ґрунти в північній частині більш
поширені.
Рельєф району не сприяє
розвитку ерозії. Відсутні великі змиви і видування верхнього гумусного
горизонту [25].
Клімат Рожищенського району
помірно-континентальний і характеризується м'якою зимою і теплим літом в
результаті впливу Атлантичного океану і материка Євразія. Середньорічна
температура повітря +7°С.
Початком весняного сезону
вважають дату стійкого переходу середньодобової температури повітря від 0°С і
вище. Весняний сезон починається з другої половини березня. Характерною
особливістю його є швидкий ріст температури, що викликає випаровування вологи
із ґрунту і насичення повітря водною парою [7].
Початок літнього сезону
вважають дату переходу середньодобової температури повітря через 15°С і вище,
яка починається в третій декаді травня. Літо не спекотне з достатнім
зволоженням.
Початком осені рахують
дату переходу середньодобової температури повітря через 10 °С до нижчої, яка
спостерігається в кінці вересня і на початку жовтня.
Міжкінцем літа і початком
осені спостерігається передосінній період, коли середньодобова температура
10-15 °С. Цей період характеризується зменшенням опадів. Погода
малохмарна, відносна вологість повітря близько 79%. В кінці жовтня
середньодобова температура досягає до +8 °С, що означає закінчення
вегетаційного періоду.
Закінчення осіннього
сезону, і початком зими вважається кінець листопада, коли середньодобова
температура повітря переходить від 0° і нижче. Тривалість зими близько 3,5
місяців до початку другої декади березня, коли температура іде вище 0°С. Зима
м'яка з невеликою кількістю опадів [7].
Глибина промерзання
ґрунтів становить 51 см з коливанням від 22 см до 89 см. Сніговий покрив
встановлюється в середньому в третій декаді грудня, а танення його починається
в кінці лютого і почату березня. Середня річна кількість опадів 618 мм.
Атмосферні опади випадають у вигляді у вигляді дошу і снігу. За весь
вегетаційний період, з опадами не менше 1 мм за добу, в середньому
нараховується 60-65 днів. Сніговий покрив найбільшої висоти досягає в лютому.
Середня висота по декадах за зиму складає 12-14 см. середній запас води в снігу
28-34 мм.
Весною, характерним є
швидкий ріст температури, за рахунок чого спостерігається інтенсивне
випаровування води із ґрунту. Але відносна вологість повітря у зв'язку з
високою температурою вдень зменшується до 66%.
В окремі роки можливі
ґрунтові і атмосферні засухи. Ґрунтові засухи бувають на піщаних ґрунтах, які
погано затримують вологу, а також коли випадає мало опадів можливі і суховії.
На території району переважають вітри західних напрямків. Зимою переважають
вітри відрогів сибірського антициклону, влітку - з Атлантичного океану. Ці
вітри влітку пом'якшують температуру, а зимою приносять відлигу.
Сильними вітрами вважають
такі вітри, коли швидкість їх перевищує 15 м/сек. Сильні вітри у весняні і
літні місяці впливають на режим зволоження, збільшується випаровування і
сприяють утворенню вітрової ерозії. Район отримує 92,7 ккал/ кв.см сумарної
сонячної радіації за рік.
Період додаткового
радіаційного балансу складає 8 місяців. Максимум спостерігається в червні 68
ккал/кв.см. Сонячні дні займають десяту частину року, половину - перемінно
хмарні, решту частину - похмурі. Найтепліпіий місяць - липень, середньодобова
температура повітря досягає + 18 °С - +19 °С. вегетаційний період станрвить 207
днів. Сума активних температур більше 10 °С складає 2840-2930 °С. більше 100
днів у році має середньодобову температуру вище +15 °С (період інтенсивної
вегетації) [25].
Сукупність природних умов
Волинського Полісся, в зоні якого розташований Рожищенський район, визначило
його ґрунтовий покрив, характерною особливістю якого є широке поширення
дерново-підзолистих ґрунтів, що є переважаючою генетичною групою з дерновими і
болотними ґрунтами [7].
Дерново-підзолисті ґрунти
займають найбільшу площу в районі (38230 га, або 47,5%). Ґрунтоутворюючою
породою для них служать воднольодовикові відклади і донні морени, а
підстилаючою - крейда, мергель і мергелізовані суглинки. Переважаючого
ґрунтоутворюючою породою тут являються водно-льодовикові відклади,
глинисто-піщаного, супіщаного механічного складу.
Дерново-підзолисті ґрунти
залягають як на підвищених слабохвилястих або вирівняних, так і понижених
елементах рельєфу (глеєваті і глеєві різновиди). Різноманітність в механічному
складі наклало помітний відбиток на їх морфологічні ознаки і властивості.
Піщані і глинисто-піщані
різновиди, особливо ті, що залягають на піщаних породах, характеризуються
великою водопроникливістю, дуже низькою вологоємністю, дуже високою ступінню
аерації. На цих ґрунтах після зволоження швидко наступає дефіцит вологи.
Водний режим супіщаних і
особливо, легкосуглинистих різновидів помітно кращий -
водопроникність їх менше, а вологоємність вище.
Глеюваті і глеєві
різновиди дерново-підзолистих ґрунтів характеризуються більш високим
зволоження. В сухі роки культурна рослинність на цих ґрунтах добре забезпечена
вологою. Разом з тим, у вологі роки на глеєвих різновидах може відмічатися
погіршення стану, а в деяких випадках і загибель посівів у зв'язку з вимоканням.
Дерново-підзолисті і тим
більше легкосуглинисті ґрунти характеризуються більш високою родючістю, ніж;
піщані і глинисто-піщані, але вони мають негативні відмінності, які характерні
для піщаних ґрунтів: малий гумусовий і орний шар, бідність на поживні елементи,
погану водоутримуючу здатність, значну кислотність.
Серед усіх
дерново-підзолистих ґрунтів, дерново-підзолисті супіщані і легкосуглинисті
ґрунти і їх глеєваті і глеєві різновиди є одними із найбільш цінних. На цих
ґрунтах залягає 17519 га ріллі, або майже 40% від загальної площі ріллі району,
а на всіх різновидах дерново-підзолистих ґрунтів залягає більше 67% ріллі
району.
Опідзолені ґрунти
представлені світлосірими, сірими, темно-сірими і чорноземами опідзоленими
супіщаними і легкосуглинистими. Залягають на підвищених елементах рельєфу.
Загальна площа опідзолених ґрунтів складає 7551 га, в тому числі світлосірі і
сірі різновиди займають 6324 га. Із загальної площі опідзолених ґрунтів 7132 га
або більше 94% їх використовується під ріллею і багаторічними насадженнями.
З агропромислової точки
зору всі опідзолені ґрунти і особливо темносірі і чорноземи, являються кращими
в районі. Потрібне вапнування.
Чорноземно-лучні ґрунти
займають незначну площу - 199 га. За даними лабораторних аналізів, ці ґрунти
мають в середньому близько 5% гумусу, рН - 6,5, тобто реакція ґрунтового
розчину близька до нейтральної.
Перегнійно-карбонатні
супіщані і легкосуглинисті ґрунти займають 2401 га. Ці ґрунти сформувались на
елювії щільних карбонатних порід і поширені окремими ділянками серед
дерново-підзолистих і опідзолених ґрунтів.
Всі перегнійно-карбонатні
ґрунти повністю насичені двохвалентними основами, особливо кальцієм, в зв'язку
з чим мають лужну реакцію ґрунтового розчину рН 7,2-7,8. Кількість гумусу
складає в середньому 2,5 % з відхиленням 1,9-3,6% в супіщаних і 3,7 з
відхиленнями 1,7-5,3 в суглинистих ґрунтах.
Дернові і лучні ґрунти
представлені піщаними, глинисто-піщаними, супіщаними і легкосуглинистими
різновидами. Ґрунтоутворюючою і підстилаючою породою є елювіальні і
воднольодовикові відклади різного механічного складу, мергелізовані суглинки і
щільні карбонатні породи Характеризуються профілем чорноземного типу. В лучних
ґрунтах гумусовий горизонт разом з перехідним складає 50 см і більше, в
дернових - менше 50 см. Відносно до сольового режиму і характеру
ґрунтоутворюючих і підстилаючих порід лугові і дернові ґрунти діляться на
типові, опідзолені і карбонатні.
Лучно-болотні ґрунти
представлені супіщаними і легкосуглинистими різновидами. Ці ґрунти мають
високий вміст гумусу і поживних речовин, але вони знаходяться у шкідливих для
рослин кислих формах. За даними лабораторних аналізів, вони містять у верхньому
шарі гумусу до 10%, рухомого азоту 18,4 мг, фосфорної кислоти 17 мг, калію
близько 5 мг на 100 г ґрунту. Реакція ґрунтового розчину слабокисла або близька
до нейтральної рН 5,5-7,0.
Ґрунти перезволожені,
мають погану аерацію і несприятливий тепловий і водний режим. Весною і осінню
затоплюються водою, в результаті чого порушується мікробіологічна діяльність.
Болотні ґрунти району
представлені болотними, торфоболотними, торфянисто-болотними і торф'яниками
низинними різної потужності.
Болотні ґрунти
характеризуються відсутністю суцільного шару торфу. Торфянисто-болотні ґрунти
мають шар торфу до 20 см, торфово-болотні від 20 до 50 см, торф'яники низинні
більше 50 см. Загальна глибина торфу торф'яників низинних в середньому складає
1,5-2 м, а в окремих місцях досягає 3 м і більше. За даними аналізів торф'яники
бувають слабо-, середньо-, добре-, і сильно розкладені. Зольність доливається
від 7 до 60 і більше процентів. Реакція ґрунтового розчину переважно слабокисла
або близька до нейтральної [7].
Рис. 1. Карта-схема Рожищанського району:
- місця збору матеріалу
РОЗДІЛ 2. МАТЕРІАЛ І МЕТОДИКА ДОСЛІДЖЕННЯ
Матеріал для дипломної
роботи був зібраний протягом 2009-2010 рр. у Рожищенському районі. Було
відібрано 120 проб. Місця відбору проб вказані на карті-схемі.
Для вивчення видового складу грунтової мезофауни
ми користувалися методиками М.С. Гілярова та К.К. Фасулатті [10; 35].
Основний етап будь-якого польового
грунтово-зоологiчного дослiдження - визначення чисельності ґрунтових тварин з
точністю, що дозволяє порівнювати чисельнiсть дослiджуваних об’єктів на рiзних
дiлянках, а також виявлення закономiрностей розподiлу органiзмiв у грунтовому
профілі.
Такі данi необхiднi для оцiнки грунтоутворюючоi
ролi грунтових тварин у рiзних її аспектах (визначення кiлькостi руйнiвникiв
рослинних залишкiв, виявлення чисельностi тварин, що прокладають у грунті ходи,
чим впливають на пористiсть, водопроникнiсть i аерацiю грунту, i т.д.).
Необхiднi вони i для визначення заселеностi грунту шкiдливими органiзмами
(личинки хрущiв i т.п.), а також чисельностi тих комах, що ушкоджують надземнi
частини рослин, але ідуть у грунт на зимівлю чи для заляльковування (капустяна
совка, соснові пильщики, гусінь лунки срiблястоi й iншi шкiдники сiльського та
лiсового господарств).
Методи облiку тварин, що живуть чи зустрiчаються
в грунті схематично можна роздiлити на прямi i непрямi.
Прямi методи - це такі, котрі дозволяють
дослiднику одержати цифри, що показують кількість об’єктів на одиницю площi
поверхні, або на одиницю об’єму ґрунту.
Непрямi методи - це такі, котрі, хоча i не дають
досить надiйного уявлення про чисельнiсть об’єкта обліку, але дозволяють
порiвнювати в бiльшiй чи меншiй мipi заселенiсть рiзних дiлянок. До непрямих
методiв можна вiднести, наприклад, облiк за плугом в борознi великих помiтних
комах (таких як личинки хрущiв) чи дощових червiв. Цей метод вимагає введення
коефiцiєнтiв перерахування, що повиннi бути для даних грунтово-клiматичних i
сезонних умов визначенi на основі порiвняння з точними даними, добутими
методами прямого облiку.
Незважаючи на привабливу легкість цього методу,
рекомендувати його можна тiльки для порiвняння даних, одержуваних в один час у
подібних по грунтових умовах мiсцевостях, i для одержання орiєнтованих цифр.
Цей метод можна використати при дослiдженнях iз сільськогосподарської
ентомологii на польових i лугових землях, але він не придатний нi в лiсi, нi в
гiрських мiсцевостях, нi в тих випадках, коли потрібен високий ступінь точностi
обліку.
Це вiдноситься i до таких методiв, як порiвняння
чисельностi шкiдливих грунтових комах на рiзних дiлянках по числу ушкоджених
рослин, тому що на шкiдливiсть личинок, що пiдгризають корені рослин, впливає
багато факторiв - вологiсть грунту, вміст у нiй гумусу, наявнiсть насiння
бур'янiв, що проростають, i т. д. Пiдрахунок грунтових личинок комах, що
концентруються на приманках, теж вiдноситься до числа вiдносних методiв.
Для облiку гyceнi i жукiв-чорнотiлок рекомендують
розкладати купки скошеної трави - це так званi концентруючi приманки. Пiд
такими приманками на дiлянках з високою щiльнiстю залягання цих шкiдникiв
скупчується iнодi по кiлька десяткiв особин пiд кожною купкою (перевiрку i
пiдрахунок варто проводити в paннi години на наступний ранок пiсля розкладки).
Але, зрозумiло, i цей метод недостатньо надiйний.
Непрямi методи можуть тiльки доповнювати й уточнювати данi, одержуванi iншими
методами, але не замiняти їx, тому що ефективнiсть непрямих методiв
визначасться занадто багатьма факторами.
Зокрема, нами були застосовані методи грунтових
розкопок, ловчих ліній для мезофауни, а також спеціальні методи обліку дрібних
членистоногих та грунтових червів [10; 35].
Метод ґрунтових розкопок
Для з'ясування якісного і
кількісного складу тварин ґрунту, перегнилого листя і трави, а також підземних
частин рослин користувалися залежно від поставлених завдань різними методами.
Найбільш поширений облік ґрунтової мезофауни метод розкопок.
Устаткування: лопата, клейонка чи щільна матерія,
кілочки (4шт.), мотузка, тканинні мішечки, банки, з 70°-ним спиртом з
додаванням декількох крапель гліцерину і формаліну, 70% розчин С2Н5ОН,
лінійка.
Виберали кілька ділянок для закладки пробних
площадок у різних частинах лісу. Розмір обраної пробної площадки визначається в
залежності від ступеня зволоженості ґрунту, найчастіше 0,25 м2 (0,5 х
0,5 м), у сухих районах до 1-2 м2 (1 м х 2 м). Чим
більше закладено ґрунтових розкопок, тим точніше проводиться виявлення видового
складу і кількості тварин. Відстань між розкопками - 5-10 м. Глибина ґрунтових
розкопок - 30-50 см, у сухих місцях на легких ґрунтах - до 100 см і більш. З
розкопки чи ґрунтового розрізу ґрунт вибирають пошарово чи по генетичних
обріях. Розкопки проводили у такий спосіб, відзнали розмір площі, забивають по
кутах кілочки, натягають між ними мотузку. Від границь відміряної площадки
відгрібали у різні сторони підстилку, оскільки проба береться в лісі чи суху
землю поверхневого шару (поряд із площадкою поміщають клейонку чи щільну
матерію, на яку потім клали вибраний з розкопки ґрунт.
Знайдених на поверхні ґрунту тварин рахували
окремо. Після приступали до викопування ґрунту лопатою. Невеликі порції ґрунту,
що викидаються на розкладену поруч з площадкою клейонку ретельно перетирають
руками, розбиваючи великі грудки, розриваючи дерновину. Весь ґрунт із шару, що
розбирається, порцію за порцією перетирають у висячому положенні між долонями,
ретельно стежачи за ґрунтом, що зсипається на клейонку, і збираючи падаючих
тварин. Тварин збирали окремо з кожної проби. Безхребетних, які потребують
спеціальної складної фіксації (дощових червів, молюсків), поміщають у тканинні
мішечки чи банки з невеликою кількістю взятої для проби ґрунту. Хижаки повинні
бути поміщені поодинці. Дрібних комах, багатоніжок, мокриць для фіксації
поміщають у посудину з 70°-ний спиртом з додаванням декількох крапель гліцерину
і формаліну, великих комах - у морилки чи посудини зі спиртом.
Зібраний і зафіксований матеріал підлягав
систематичній обробці для визначення видової приналежності для чого
використовували спеціальні визначники, якими користуються для ідентифікації.
Облік дрібних членистоногих
Для обліку дрібних членистоногих проби можна
брати на світлозахищеній стінці чи траншеї ґрунтового розрізу методом змішаних
зразків з кожного генетичного шару або суцільним стовпчиком, розділеної на
окремі зразки на глибину ґрунтового профілю від at
до В і. В останньому випадку використовується рамка 5х5 см. Кожна серія
зразків являє собою суцільну колонку ґрунту на глибину до 40 см, розділену на
частини, що відповідають генетичним шарам. При товщині шару, що перевищує
висоту рамки (5 см), шари поділяють на кілька послідовних зразків. У нормальній
непорушеній частині ґрунтового профілю природна границя вертикального розподілу
мікроартропод - підзолистий обрій, на глибині якого знаходиться лише 3%
загального населення. У випадку методу змішаних зразків досліджується погонний
метр траншеї, розділений на 5 колонок. У кожнім зі стовпчиків із середини
ґрунтових шарів береться дрібним
буром по 5 точкових зразків, об'єднаних потім в одну змішану пробу обсягом
150см3 (обсяг бура 30 см3).
Таким чином, протягом досліджуваного ґрунтового
метра траншеї з кожного генетичного обрію береться по 5 змішаних зразків. З
підстилки беруть по 5 об'ємних зразків і з'єднують їх в один, обсяг бури
відповідає кількості підстилки на площадці 3 х 3 см. Чисельність
тварин виражають числом екземплярів особин у шарі ґрунту товщиною 1 м.
Метод ловчих ліній для мезофауни
Для обліку мезофауни використовується також
стандартний метод ловчих ліній. Ця методика широко застосовується для збору
активних на поверхні ґрунту комах, наприклад, жужелиць, жуків і ін. Лінії
виставляються двома рівнобіжними рядами з визначеними проміжками між пастками
Барбера (до 10 м). У лінії може бути від 5 до 10 пасток.
Устаткування: скляні банки ємністю 0,5 л з
діаметром 7,5 см, 4%-ний розчин формаліну, кришки з пластмаси розміром 14 х
14 см (установлюють над пастками на висоті 4-5 см для захисту від дощової
води).
На дно банок поміщають невелику кількість ґрунту
для створення сприятливих умов комасі, що туди потрапила. Пастку ставлять від
декількох діб до 10 діб. При цьому вибір комах виражається в одиницях
динамічної щільності - вловимістю 10 ловушок/добу. Облік загальної кількості
тварин, і так само домінантних і субдомінантних видів. Домінантними вважають
види, чисельність яких складає більше 5%, субдомінантами - 2-5% від загального
числа зібраних особин.
Облік ґрунтових червів
Для обліку енхитреїд застосовуються спеціальні
методи Нільсена та Оконнора.
1. Метод Нільсена, який заснований на принципі
створення температурного градієнта. Вимагає наступного устаткування:
циліндричні металеві посудини (можна використовувати консервні банки діаметром
близько 20 см, з дірчастим дном), гравій, дротове сито, судина з водою.
На дно циліндричної металевої посудини насипають
шар гравію висотою близько 3 см, вище якого вставляють щільно притиснуте до
стінки дротове сито. На сито поміщають пробу ґрунту, зверху присипають її
вологим піском. Посудини з пробами вставляють у посудину з водою,що
нагрівається на повільному вогні, налитої до висоти гравію в судинах із
пробами. Нагрівають воду до 60-65° і витримують близько 2 год. За цей час черви
виповзають з більш гарячого субстрату в пісок, звідки їх витягають і
підраховують.
2. Метод Оконнора.
Устаткування: велика лійка (діаметр близько 10
см), гумова трубку з затиском, сито, 60-ватна електрична лампочка, годинник.
На носик лійки надягають гумову трубку з
затиском, під якою підставляють посудину. У лійку наливають доверху воду. У
верхній частині лійки зміцнюють сито, на якому розподіляють пробу ґрунту так,
щоб ґрунт виявився зануреним у воду. Над пробою включають електричну лампочку.
При нагріванні проби енхитреїди мігрують вниз і, проповзаючи крізь осередки
сита, тонуть у воді, накопичуючи в носику лійки й у гумовій трубці. Після 3
годинної вигонки відкривають затиск і хробаки зі струменем води попадають у
поставлену судину, після чого їх легко підрахувати.
Обидва методи дають близькі величини при
дослідженні мінеральних ґрунтів. При вигонці з підстилки другий метод дозволяє
вирахувати в 1,5 рази більше червів, ніж перший. Дослідження рекомендується
проводити кожен місяць у плині теплого сезону року.
Біоценометри
Для кількісного підрахунку безхребетних на
поверхні землі, в травостоях, кущах використовували біоценометри. Вони бувають
різних систем, але всі вони побудовані за одним принципом. Визначену ділянку
землі в 1 або 0,25 м2, накривали пристроєм, потім з нього вибирали
всіх тварин, підраховували їх, і вираховували на визначену площу.
Біоценометр Баскіной і Фрідмана. Являє собою
квадратний залізний ящик з двома ручками. Нижній край ящика загострений , а
верхній загнутий назовні для надягання мішка. Мішок зшитий із полотна по
розміру ящика. Перед початком роботи з приладом мішок надягали на ящик і
зав’язували. Біоценометр брали за ручки і піднімали вгору на витягнуті руки і
швидко опускали на землю, потім землею навколо підкопували лопатою, ділянку
землі піднімали і швидко підштовхували в мішок. Мішок знімали і відразу ж
зав’язували [35].
Термоеклектор (апарат Тульгрена )
Широке застосування знайшли електричні
термоеклектори для підрахунку комах лісової підстилки, для відбору тварин із
грунту. Розрізняють термоеклектори одинарні, трьохгніздні і багатогніздні, але
принцип роботи у всіх один, він полягає у взаємному впливі температури, світла
на мешканців досліджуваного субстрату. Тепловий і світловий таксис заставляють
членистоногих переходити із добре освітлених і нагрітих шарів грунту вниз,
звідки вони через сітку еклектора скочуються по нахиленій поверхні отвору в
посудину з фіксатором, яким може бути 70% спирт, рідше
використовують воду.[35]
Обробка та фіксація
матеріалу
Зібраний матеріал
фіксували кип'ятком - спосіб швидкої фіксації і застосовували повільну фіксацію
в хромовій суміші: формаліну (40%) - 80 частин за об'ємом, розчину хромової
кислоти у воді (1%) - 180 частин, оцтової кислоти (крижаний) - 10 частин. У
хромовій суміші матеріал потрібно тримати декілька днів (але не більше місяця),
потім промити і перенести в консервуючу рідину. Личинки крупних комах
(пластинчатовусих) в цій суміші фіксуються погано. Для них краще застосовувати
кип'яток. У посудину, де знаходяться личинки, наливали кип'яток, варили 2-3 хв.
на повільному вогні, після чого давали остигнути. Потім личинок поміщали в 96%
спирту, а через декілька днів переносили їх в спирт меншої (70%) концентрації і
в ньому зберігали. Спирт можна замінити сумішшю Мак-Грегора (10 см3)
нерозведеного формаліну, 10 смсм3 води, що дистилює, і 2 см3
гліцерину). При фіксації на тілі крупних м'яких личинок рекомендується зробити
декілька надколів голкою, що прискорює просочення тканин фіксуючою рідиною.
Добре фіксуються личинки і без нагрівання рідиною наступного складу:
концентрований формалін - 5 - 6 частин, етиловий спирт (96%), - 15 частин,
крижана оцтова кислота-2 частини, вода, що дистилює - 30 частин. Через 10 днів
личинок слід перенести в 80% спирту, який через декілька днів слід змінити. Для
фіксації дрібних м'яких личинок користувалися рідиною такого складу: спирт -
96% - 750 частин, сірчаний ефір - 250 частин, крижана оцтова кислота - 30
частин, формалін 40%-ний - 3 частини. Зберігали личинок в пробірках в темній
шафі, оскільки світло обезбарвлює і розм'якшує фіксованих в спирті личинок. У
кожній пробірці обов'язково повинна бути етикетка, яка пишеться на кальці або
щільному папері графітним олівцем або тушшю. Етикетки, написані тушшю,
рекомендується перед опусканням в спирт підігріти над полум'ям, а краще
змастити розчином целоїдину в ацетоні.
Для збереження матеріалу
застосовували консервацію суху або в рідинах. Суха консервація полягає в
наступному: личинку комахи поміщають в посуд або коробку і засипають дрібно
товченою сіллю. Цей спосіб не завжди дає добрі результати і тому має обмежене
застосування.
Для консервації в рідині найчастіше
використовували 2-5% формалін або 70% спирт. [35]
Рідко доводиться обробляти матеріал паралельно з
розкопками. В більшості випадків його обробляли пізніше - в зручнішій
обстановці тому всі збори необхідно зафіксувати і детально етикетувати.
Рекомендується така форма етикетки:
№____Місце______Час_________Характер
місцевості____________
Спосіб взяття
спроби_______________
|
Назва
тварини
|
1 шар
|
2 шар
|
3 шар
|
4 шар
|
|
Глибина
кожного шару
|
|
5 см
|
10 см
|
20 см
|
30 см
|
|
|
|
|
|
РОЗДІЛ 3. АНАЛІЗ РЕЗУЛЬТАТІВ ДОСЛІДЖЕННЯ
.1 Еколого-фауністична
характеристика грунтової
Мезофауни Рожищенського району
Мезофауна - об’єднує досить різноманітну і
багаточисельну групу грунтових тварин середніх розмірів. Основну масу мезофауни
становлять грунтові олігохети родини люмбриціди, багатоніжки, комахи та їх
личинки.
Екологічні групи ґрунтових тварин виділяються не
тільки за розмірами, що відображає безпосередньо їх вплив на ґрунт, але також і
за типами живлення, що визначає положення організмів у трофічних ланцюгах
біотичного співтовариства. Серед ґрунтових тварин є наступні трофічні групи:
сапрофаги, фітофаги, зоофаги, некрофаги.
Сапрофаги - найбільш численна й важлива за
значенням група ґрунтових тварин. Вони переробляють мертві залишки рослин, опад
і відпад як на поверхні ґрунту - у підстилці, так і в зоні кореневих систем. До
них належать черви, багатоніжки, мокриці, деякі кліщі й личинки комах. Це
екологічне угруповання становить найбільший інтерес щодо вивчення ролі тварин у
перетворенні органічних речовин у ґрунті. До сапрофагів відносять:
копрофагів, що живляться екскрементами інших
тварин, головним чином, ссавців;
детритофагів, що використовують у їжу детрит -
дрібні органічні частки разом із живими мікроорганізмами, що їх населяють і
розкладають.
Фітофаги харчуються тканинами коренів живих
рослин, завдають збитки сільському господарству. Наприклад, личинка травневого
хруща підгризає корені молодих сіянців сосни. Різновидом фітофагів є альгофаги,
що харчуються водоростями.
3оофаги поїдають інших тварин, виступаючи в ролі
хижаків або паразитів. Прикладами можуть бути всі комахоїдні тварини, що
живляться найпростішими і коловертками, хижі кліщі, що нападають на нематод,
коллембол, енхітреїд.
Некрофаги використовують у їжу трупи тварин,
виступаючи в ролі санітарів у природних екосистемах.
Педобіонти мезофауни розділені також на три
групи:
геобіонти - постійні мешканці ґрунтів (наприклад,
дощові черви, багатоніжки, ногохвістки),
геофіли, що живуть у ґрунті протягом частини
життєвого циклу (личинки хрущів, коваликів),
геоксени, що лише тимчасово мешкають у ґрунті.
Усі вони становлять складну систему, що одержала
назву сапрофільного комплексу тварин.
Ґрунтові олігохети родини люмбриціди
На основі опрацювання літературних джерел та
аналізу результатів власних досліджень було встановлено, що в складі
олігохетофауни цього регіону зараз нараховується 14 видів люмбріцід, які належать
до 8 родів.octaedra (Savigny, 1926)
В біоценозах району дослідження є одним з
поширених видів. Належить до підстилкової морфо-екологічної групи. В.К. Балуєв
відносить його до верхньоярусної морфо-екологічної групи.octaedra поширена у
листяних, мішаних і хвойних лісах, на луках. Особини виду живуть у різних
біотопах лісів, а саме: у підстилці, гниючих пеньках, стовбурах, гілках дерев.
Dendrobaena octaedra знаходили і у верхньому горизонті лісових. Найвища
чисельність його відмічена у біоценозах листяного, хвойного і мішаного лісів,
луках.octaedra є евритопним видом і має здатність заселяти біотопи з різною
вологістю ґрунтів: вологі, сирі та мокрі. Найвища чисельність зафіксована у
мокрих і сирих біотопах.
Особливістю виду є його гігрофільність та
приуроченість до лісової підстилки. За несприятливих умов черви можуть
знаходитись у верхньому шарі ґрунту на глибині до 7-9 см. Найчастіше, під час
міграції, заселяють нірки ґрунтово-підстилкових і нірникових морфо-екологічних
груп люмбріцід. Від розвитку мережі ґрунтових ходів залежить і їх вертикальна
міграція у просторі ґрунтового профілю. Черви дуже рухливі.rubidus
subrubicundus (Eisen, 1874)
В біоценозах району дослідження цей вид є
рідкісним і нечисленним Dendrodrilus rubidus sudrubicundus виявлений нами у біотопах
з великим нагромадженням рослинних решток (волога підстилка, гнойові кагати,
нагромадження соломи, листя, органічне сміття).
Як усі підстилкові черви, форма також поширена у
вологих і сирих біоценозах, але уникає відкритих ділянок, які швидко втрачають
вологість верхнього шару підстилки. Черви надають перевагу ґрунтам з легким
гранулометричним складом і заслуговують уваги як гумусоутворювачі.
Незначна поширеність видів у районі дослідження
районі зумовлена бідною трофічною базою, тобто малим нагромадженням органічних
решток. В цьому значну роль відіграють легкі ґрунти району дослідження, що
сприяють швидкій мінералізації органічних решток та відносно нестійкому водному
режиму.rubidus tenuis (Eisen, 1874)
Малопоширений та нечисленний вид, який заселяє рівнинні
ділянки. Звичайний мешканець біотопів із значним нагромадженням органічних
решток. Dendrodrilus rubidus tenuis поширений у найрізноманітніших середовищах
листяних, мішаних, рідше у хвойних лісах. Рідкісним є у ґрунтах лук. Можна
вважати, що вид є типовим мешканцем лісових екосистем. Його відносять до
підстилкової морфо-екологічної групи. Він заселяє не лише підстилковий
горизонт, утворений лісовою рослинністю, але й верхній ґрунтовий горизонт.
Трофічною базою виду слугує підстилка листяних лісів, трухляві стовбури дерев,
пеньки, органічні рештки стовбурів дерев, рідше поверхневий шар ґрунту з
великим вмістом коріння трав’яних рослин лук, парків. Dendrodrilus rubidus
tenuis живиться деревними, рідко трав’яними рештками і є важливим фактором
гумусоутворення на лісосіках та старих лісових насадженнях. Зазначимо, що він
поширений і на луках, пасовищах, рідше на господарських ґрунтах, які були
утворені на місці лісових насаджень. Найвища чисельність цього виду відмічена
на торф’яно-перегнійних ґрунтах вільшняків і становила 197,1 екз/м2.
Гранулометричний склад ґрунтів до виду має
опосередковане відношення, оскільки екологічною нішею D. rubidus tenuis є
підстилка. Гранулометричний склад ґрунту відіграє роль середовища, що утримує
вологу. Найбільша кількість біотопів знаходиться на глинистих ґрунтах, менше на
піщаних і суглинкових.rubidus rubidus (Savigny, 1826)
Як і попередні два види, D. rubidus rubidus
належить до підстилкової морфо-екологічної групи. Його можна віднести до
еврибіонтних організмів, які заселяють рівнинні ділянки. Серед видів роду
Dendrodrilus у Рожищенському районі зустрічається найчастіше.
Найвища чисельність виду відмічена у листяному
лісі та на луках, дещо менша - у хвойному і мішаному лісах.
Вид особливо вибагливий до вологості середовища.
Оптимальним для них є сирий ґрунт, а у вологому середовищі чисельність
люмбріцід знижується майже у два рази. У сухому середовищі представники виду
відсутні.fetida (Savigny, 1826)
Вид найчастіше трапляється біля господарських
будівель у гниючому смітті, компостах. Нами неодноразово було відмічено
знаходження олігохет у гниючих стовбурах дерев, пнях, що були попередньо
піддані дії гіфів грибів. Такі біотопи виділяються високою відносною вологістю.
Численною може бути E. fetida на корі дерев, у заглибинах, які утворилися в
місцях розходження гілок.
З усіх підстилкових дощових червів E. fetida
найенергійніше утилізує органічні рештки. З метою утилізації органічних решток
(гною, компостів, сміття населених пунктів, відходів промислових підприємств)
та отримання гумусу його розмножують у промислових масштабах. Вид відносять до
синантропних організмів.tetraedra tetradra (Savigny, 1826)tetraedra tetradra
належить до підстилкової морфо-екологічної групи. Він є одним із
найвологолюбивіших видів і його відносять до підгрупи амфібіотичних організмів.
Вид дуже поширений у болотних купинах серед коріння рослин, у гниючих пеньках,
гниючій підстилці, детриті, придорожніх ровах.
У районі дослідження E. tetraedra tetradra
заселяє заболочені ділянки заплав, береги ставків, струмків, болота, торфовища,
підмоклі пасовища і луки, гниючу мокру підстилку листяних лісів.
Значну роль у житті даних організмів відіграє
гранулометричний склад ґрунтів біотопів. Зокрема, найсприятливіші для поселення
є глинисті ґрунти, дещо менше суглинкові і супіщані.
Найвища чисельність люмбріцід відмічена у
листяній підстилці дерново-підзолистих глейових ґрунтів. В екстремальних умовах
E. tetraedra tetraedra може здійснювати міграцію в ґрунтовому профілі на
глибину до 40 см у разі наявності вертикальних нірок.
Allolobophora chlorotica
(Savigny, 1826)
В біоценозах району
дослідження Allolobophora chlorotica зустрічається рідко. Цей вид ми відносимо
до середньоярусної нірникової морфо-екологічної групи. Нерідко в представників
цього виду проявляється екологічна різноманітність, пов'язана з відмінностями в
забарвленні.
Досліджуваний вид відносять до амфібіотичних. Їм
властива найвища зустрічальність у сирих, дещо менша - у вологих ґрунтах.
Тобто, даний вид є дуже вимогливим до вологості середовища існування.
Типовими біоценозами для А. chlorotica є луки,
поля. Найвища чисельність виявлена в біотопах лук. Не зустрічається даний вид у
біоценозах лісів.caliginosa caliginosa (Savigny, 1826)caliginosa caliginosa є
еврибіонтним. Цей вид є типовим для добре оброблюваних грунтів, луків, лісів,
тощо. За дослідженням Зражевського кількість їх становить 195,7% на м2.
Найчастіше вони виявляються на іригаційних систем.
Дану форму розглядають як
2 члени поліплоїдного ряду, а саме: три- і тетраплоїди з хромосомчим числом відповідно
54 і 72. Ці форми морфологічно не відрізняються. Поліпоїди форми -
грунтово-підстилкові. Можна припустити, що в процесі адаптаціогенезу вони
перейшли в підстилковий шар і не втратили зв'язку з попередньою екологічною
нішею.
Вид поширений у ґрунтах з різним гранулометричним
складом. Надають перевагу легким (супіщані, легкі і середні суглинкові)
грунтам. roseа rosea (Savigny, 1826)
Один з найбільш поширених
серед люмбріцід. Пластичний вид.
Представники
зустрічаються в верхньому шарі грунту, на глибині близько 20 см. В нижніх шарах
можна побачити при засусі. Можлива діапауза. Це представники середньо ярусного
морфо-екологічного типу, які живляться ґрунтовим перегноєм,. Зустрічаються в
суглинах та глинах. Найчисельніші в підзолистих (бурих) грунтах. Гігрофільні
види - наявний в сирих і вологих грунтах.
Lumbricus terrestris Linnaeus, 1758
Серед представників роду люмбрікус вид є
найбільшим. Його часто використовують як об’єкт при вивченні морфо-анатомічної
будови цієї групи ґрунтових олігохет.
Рідко заселяють хвойні ліси. А.І. Зражевський
вказує на відсутність селективності виду до листяної підстилки дерев. Вид
уникає лише болотних і торфових ґрунтів, у яких відбуваються глейові процеси.
Особливістю червів цього виду, який належить до
нірникової морфо-екологічної групи, є здатність прокладати глибокі вертикальні
і горизонтальні ходи в ґрунтовому профілі, завдяки добре розвинутому
шкірно-м’язовому мішку.
Живляться особини в нічний період на поверхні
ґрунту, а протягом дня знаходяться в горизонтах ґрунтового профілю Н, НЕ, де
живляться диспергованими органічними рештками і прокладають вертикальні
ходи.rubellus Hoffmeister, 1843
В регіоні
дослідження серед представників роду люмбрікус вид є найпоширенішим. Lumbricus
rubellus поширений у мішаному лісі, рідше у хвойному. Р. Бальтцер вказує на
його домінування в кислих ґрунтах, які зайняті хвойними лісами, де він відіграє
важливу роль в утворенні гумусу. Його чисельність є найвищою в лісових
біоценозах. Має місце поширення виду і в агроценозах.rubellus здатний жити в
грубому гумусі (мор) міцелярного походження, який утворився в процесі
трансформації хвойної підстилки. Необхідно зазначити, що перебувати в такому
кислому середовищі (рН до 3,2) черви можуть лише завдяки добре розвинутим
вапняковим залозам.
Живляться ці олігохети переважно рослинними
рештками та екскрементами безхребетних тварин, які живуть на поверхні ґрунту.
За морфо-екологічним типом L. rubellus потрібно
віднести до ґрунтово-підстилкових. Особини виду тримаються переважно у
гумусовому горизонті (0-10 см) ґрунту і лише під час посухи мігрують на глибину
до 5-12 см, де знаходяться у в’ялому стані і не переходять до стану
діапаузи.baicalensis Michaelsen, 1900
Вид належить до морфо-екологічної підстилкової
групи. Черви дуже рухливі. Типовими біотопами виду є листяні ліси, підмоклі
луки, береги великих водосховищ, озер, заплавних боліт. Найвища чисельність у
листяних лісах (вільшняках).baicalensis близький до амфібіотичної групи і
заселяє біотопи із сирими і мокрими ґрунтами. Серед ґрунтів олігохети надають
перевагу мулисто-піщаним, рідше глинисто-піщаним ґрунтам. Іноді заселяють
прибережні ділянки водойм, які межують із зоною ріпалі.
Ці черви відіграють важливу роль у розпушуванні
верхнього горизонту ґрунту заплав, особливо після затоплення їх весняними та
осінніми мулистими паводковими водами. Завдяки життєдіяльності цих червів після
сходження води ґрунт швидко набуває сприятливого гідро-повітряного
режиму.transpadanus (Rosа, 1884)
Заселяє луки, листяні ліси, береги річок і
ставів. Octodrilus transpadanus рідко трапляється у мішаних і відсутній у
хвойних лісах та на оброблюваних ґрунтах. Вид належить до нірникової
морфо-екологічної групи.
Біотопи виду приурочені до ґрунтів, у яких
гранулометричний склад є середнім і важким, тобто представлений суглинками або
глинами. Проте він надає перевагу глинам. Дуже рідко особини виду трапляються у
супіщаному ґрунті. Нами виявлена найбільша щільність особин виду у лучних
ґрунтах, яка сягала 137 екз/м2. Цей вид, як і попередній, є типовим
кальцієфілом. Вони поширені у біотопах, які сформувались на крейдяних,
мергельних або вапнякових відкладах. Необхідно зауважити, що нами виявлені
олігохети цього виду у ґрунтах, де материнська порода залягала близько до
поверхні ґрунту (60-80 см). Такі ґрунти, як правило, нейтральні, слабо лужні,
рідше - лужні. Проведених аналіз проб люмбріцід, зібраних у біотопах, які
сформувались на крейдяних відкладах, вказує на домінування Octodrilus
transpadanus. У пробах особини виду становлять 97-100 %, іноді в них
трапляється Octolasion lacteum (до 3 %).lacteum (Öerley, 1855)
Один з домінуючих видів у біоценозах районі
дослідження. Заселяє більшість біотопів регіону дослідження. Типовими біотопами
виду є вологі луки, волога підстилка листяних лісів, гниючі дерева, мохи
листяних і мішаних лісів, береги рік, струмків, озер, водосховищ. Значне
поширення виду І. А. Четиркіна пов’язує із алювіальними відкладами річкових
долин. П.С. Свєтлов відзначає, що вид типовий для лісових насаджень і в пробах
становить 35,7 %.
Т. С. Перель відносить цей вид до верхньоярусної
морфо-екологічної групи. Необхідно азначити, що вид є типовим кальцієфілом і
заселяє ґрунти, які сформувались на крейдяних, мергельних і вапнякових
відкладах. Величина актуальної кислотності ґрунтів становить 7,5-8,2 і
зумовлена високою концентрацією катіонів Са2+.
За умов свіжого, вологого, сирого ґрунту з легким
гранулометричним складом О. lacteum є одним з важливих гумусоутворювачів серед
люмбріцід і бере активну участь в ґрунтоутворенні та міграції хімічних
елементів у межах ґрунтового профілю.
Ґрунтові молюски
В наземних екосистемах
регіону дослідження наявні молюски з ряду легеневих класу черевоногих. Значна
їх частина є сапрофагами і пов'язана головним чином з підстилкою та іншими
рослинними рештками. Зокрема до споживання листового опаду разом з розвиненими
тут грибами, первинними його розкладачами, найбільше спеціалізовані дрібні та
середні равлики, що більшою мірою воліють до лісової підстилки.
У роті в молюска є
м'язистий язик, покритий твердими зубчиками. Час від часу молюск висуває язик,
зіскрібає ним, як теркою, м'які частини рослин і ковтає їх. Через глотку та
стравохід їжа надходить у шлунок, а потім - у кишку. Кишка петлеподібно
вигинається всередині тулуба й закінчується на його правому боці анальним
отвором. На відміну від усіх нижчих тварин, молюск має печінку - травну залозу,
клітини якої виробляють травний сік. Отже, травна система молюсків складніша,
ніж у дощового черв'яка.
Це ледь не єдині тварини,
принаймні серед наземних, які виробляють власний фермент целюлазу. Для них взагалі
характерний високий рівень активності ферментів, котрих в гомогенаті їх
травного тракту виявлено більше 30, головним чином карбогідрази,
спеціалізованих на розщепленні вуглеводнів. Особливо важливою з точки зору
розкладання рослинних решток є здатність молюсків розщеплювати лігнін і пектин,
що дозволяє харчуватись не лише листовим опалом, але навіть гнилою деревиною.
Цінним є також те, що
крім ферментів у їх травному тракті виробляється велика кількість
мукополісахаридів, які скріплюють частинки неперетравленої їжі, завдяки чому
екскременти мають велику механічну міцність і довго зберігаються, в них
розвивається процес гуміфікації, який розпочався ще в травному тракті. За цими
рисами вони подібні до копролітів дощових червів, які як і останні так само істотно
поліпшують структуру ґрунту, формують дрібнозернистий поверхневий його шар
мулового типу, збагачують корисними органічними та мінеральними речовинами.
Для молюсків, що харчуються листовим опадом
властива певна вибірковість. Середній рівень споживання по відношенню до маси
(що особливо істотно, оскільки значна частина загальної ваги припадає на
черепашку) коливається у різних видів в межах 0,5 - 5,2 %, а у перерахунку на
суху вагу в середньому складає 16% на добу. Засвоєння різних видів опаду
становить від 43 до 58 % причому ця величина прямо пропорційна вмісту
клітковини. В рівнинних лісах Європи їх питома чисельність сягає близько 500
особин, а біомаса 278 г на м. При такій щільності поселення молюски можуть
переробити за сезон 35-43% річного опаду і отже є активними учасниками його
розкладання.
Особливу групу наземних молюсків становлять голі
слизні їх черепашка повністю обростає мантією і стає рудиментарною, а в деяких
повністю зникає. Слизні живуть в тих місцях, де достатня вологість. Головна
зона їх розселення ліс. Їжею служить свіжоопавші листки, відмерлі трави. Вони
попадають на живі рослини, але тільки при нестачі іншої їжі. Деякі з них
живляться слизнями, червами.
Найбільш поширеним є польовий слизень
(Agriolimax agrestis) - довжина 35 - 40 мм, мантія займає до 1/3 довжини тіла.
Забарвлення від майже білого до кремового, без темного малюнку. Весною він
нападає на сходи зернових культур. В роки масового розмноження наносить шкоду
для господарства.
Великий слизень (Limax maximus), довжина
тіла до 130 мм. Мантія займає 1/3 довжини тіла. Забарвлення строкате на жовтому
чи білому фоні 2-3 пари темних полосок. Щупальця однокольорові, без темних
цяток. Наносить шкоду деревним породам.
Слимак чагарниковий (Bradybaena fruticum).
У цього слимака майже гладка шароподібна черепашка, висотою 16-17 мм, шириною
18-20 мм з 5-6 оборотами. Забарвлення можк бути різним, від сірувато - білого
до червоного забарвлення.
Садовий слимак (Cepae hortensis). У
садового слимака черепашка клубоподібна, схожа на черепашку попереднього
слимака. Висота черепашки 15 - 16 мм, ширина 18 - 21 мм з 4 - 5 оборотами, на
всіх оборотах видніються темні спіральні полоски.
Янтарка звичайна (Succinea putris). Назву
свою отримала за яскраво - жовтий колір, витягнутою, тонкою, ламкою черепашкою.
Висота її 16 - 22 мм, ширина 8 - 11 мм. Ракушка з 3 - 4 оборотами, останній
оборот сильно видутий і розширений. Мешкає в сирих місцях.
Багатоніжки (Myriapoda)
Багатоніжки живуть, в основному, на землі
або в самих верхніх шарах грунту. Два найбільш відомі представники - кістянка
(Lithobius) і ківсяк (Julus). При рихлені грунту лісу, в парку, саду і при коні
ям можуть зустрічатися як ці дві форми, так і різні інші багатоніжки.
Досить часто трапляють в грунті дуже тонкі
жовті чи червоно- жовті багатоніжки із родини Geophilsdae. Потривожений
Geophilus починає відразу повзати по руці чи папері, де добре помітно головку
без очей, з парою добре помітних ниткоподібних вусиків і багаточисельні ноги
(19 - 45 пар). Це хижак, який живиться дрібними комахами, червами. Молоді
Geophilus по всьому схожі на дорослих, але відрізняються більш блідим
забарвленням тіла.
З другого ряду багатоніжок можна зустріти
звичайного Polydesmus complanatus - невелику сліпу багатоніжку довжиною до 2 см
коричнево-сірого кольору з плоским тілом із видніючимися з боків краями спинних
щитиків, яких налічується до 20. На голові знаходиться пара невеликих вусиків,
число ніжок доходить до 38. Як і інші багатоніжки має пахучі залози, які
виділяють секрет зі специфічним, який нагадує синильну кислоту запахом, що
з’являється після доторкання пальцем до тварин. Живиться Poludesmus
підгниваючими рослинними рештками, інколи може шкодити сходженню насіння,
під’їдаючи їх корінці. Трапляється і знаходити гніздо, які він робить із
частинок землі у вигляді невеликого дзвону з отвором для повітря. В ньому
знаходиться кучка білих яєць, з яких виходить молодь з трьома парами ніжок.
Ще один представник багатоніжок з ряду
Diplopoda - Polyzonim germanicum Brandt, який зустрічаєть в тінистих сируватих
місцях. Це невелика тварина, довжиною до 15 мм з випуклим зверху і знизу тілом,
гладкою блискучою поверхнею шкіри, невеликою головою, витягнутою вперед у
вигляді дзьоба, зверху непомітною, з маленькими вусиками і багаточисельними до
90 пар ніжками. Його можна знаходити згорнутим в будь - якому заглиблені
грунту. Багатоніжка повзає по руці досить швидко. При доторканні з’являється
різкий запах камфори. На початку літа можна знайти в землі і її і
яйця,відкладені купкою, навколо якої тримається самка. Із яєць виходить молодь,
яка має, на відміну від інших 4 пари ніжок, число яких потім, як зазвичай
збільшується [10, 62 ].
Комахи (Insecta)
Ковалики (Elateridae)
Личинки жуків-коваликів
(дротянки) живуть у ґрунті. У коваликів голова личинки зверху плоска, верхня
губа не видніється із під головного щита, вусики короткі, не видаються на
передній край голови, всі ноги однакової величини.
Різні види розрізняються по будові
заднього кінця тіла, який може бути закрученим або тупим, або роздвоюється з
двома відростками які у різних видів відрізняються за формою.
Розвиток у коваликів продовжується до
п’яти років. Оляльковування проходить завжди в землі. Дорослі жуки ковалики
зустрічаються на різних рослинах, можуть об’їдали листки і квіти, тоді як
чорнотілки тримаються зазвичай на землі.
За типом харчування
відповідні групи дротянок варто назвати так:
) фітофаги; 2)
фітозоофаги; 3) зоофітофаги: 4) хижаки; 5) облігатні хижаки.
За типом харчування В.Г.
Долін [11] поділяє дротянок на наступні групи.
1.Усеїдні, але переважно
фітофаги, пов'язані зі злаковою рослинністю (роди Agriotes, Ectinus).
Нормальний розвиток личинок може відбуватися і без споживання тваринної їжі.
2.Усеїдні, із сильно
вираженою фітофагією, але потребують для нормального розвитку їжу тваринного
походження (роди Corymbites, Selatosomus).
3.Усеїдні, зі слабко
вираженою фітофагією, які надають перевагу рослинним залишкам І хижацтву (роди
Melanotus, Limonius, Athous). При дефіциті вологи в грунті або відсутності
тваринної їжі личинки цієї групи серйозно пошкоджують насіння, підземні стебла
і коріння-бульбоплоди.
4.Хижаки і некрофаги,
здатні до сапрофаги, що не пошкоджують живі рослини (роди Cardiophorini,
Elater, Synaptus, Prostemon, деякі підроди роду Athous).
5.Облігатні хижаки (роди
Lacon, Adelocera, Hypoganus, Calambus і підроди роду Athous: Diacanthous,
Stenagostus).
У досліджуваному регіоні типовими
представниками жуків-коваликів є:
Ковалик буроногий (М. rrunipes) - тіло не
дуже витягнуте, надкрила з добре розвинутими боріздками в передній частині.
Останнє черевне кільце із двома вдавлюваннями, на вершині обрубане, посередині
мохнате. Чорний в жовтуватих волосках, вусики і ноги темно-бурі. Довжина тіла
12-17 мм. Личинки довжиною до 19 мм, розвиваються в гнилій деревині, їдять
личинки і лялечки других комах.
Ковалик кривавий (Elater sangyines)
довжина тіла 9-12 мм предспинка темна на яскраво чорних надкрилах в середині
поздовж знаходиться чорна пляма. Активний в травні-червні. Личинки довжиною до
22 мм, розвиваються в гнилій деревині, їдять личинки і лялечки других комах.
Личинки жовтувато-коричневі.
Ковалик чорний (Athous niger) довжиною 10
- 14мм тіло зверху чорне, надкрила бурі в не густих сірих волосках. Личинки
всеїдні і полюбляють хижаків.
Ковалик червононогий (М. rufipe) - тіло
сильно витягнуте; надкрила в 3 - 4 рази довші передньоспинки, їх повздовжні
боріздки тонкі. В передній частині погано розрізняються. Щиток набагато довший
від своєї ширини. Чорний або буро-чорний, вусики і ноги червоно-бурі. Личинки
темно-коричневі.
Пластинчастовусі (Scarabidae)
Усі личинки
пластинчастовусих живуть у грунті або в субстратах, близьких до грунту за
механічними властивостями (гниюча деревина, що розкладається, рослинні тканини
і речовини тваринного походження, екскременти хребетних тварин). Личинок, що
відкрито живуть, серед представників пластинчастовусих немає. У зв'язку з цим
спостерігається відносна одноманітність форм личинок, будова яких зводиться до
деяких типів, які не сильно розрізняються, на відміну від дуже різноманітних
личинок тих родин жуків, де є великі відмінності в середовищі існування, як,
наприклад, у листоїдів (родини Chrysomelidae).
Питання про адаптивне
значення форми личинок комах, що живуть у ґрунті і подібних субстратах,
детально розроблене М.С. Гіляровим [10]. Загальні висновки його такі: С-подібна
форма тіла личинок (такими є личинки пластинчастовусих) є адаптацією до
активного руху в специфічному, дуже важкому за механічними властивостями
середовищі, яким є ґрунт або щільна рослинна тканина.
Спосіб пересування
личинок пластинчастовусих у грунті також вивчений і описаний М.С. Гіляровим
[10].
За способом життя і
харчування серед личинок виділяються наступні групи:
1.Личинки, що вільно живуть у ґрунті й
активно розшукують їжу, яка складається з коренів (узагалі підземних частин)
рослин, переважно живих, але також і відмерлих. Ці личинки, особливо в молодому
віці, значною мірою харчуються і перегноєм. Сюди відносяться личинки хрущів із
триб Ratelini, Melolonthini, Rhizotrogini, Pachydemini, Sericini, Hopliini,
Glaphyrini і, частково, Dynastini (Pentadon). Значна частина личинок цієї групи
є шкідниками сільськогосподарських і лісових культур. Багато видів шкодять і в
імагінальній фазі.
2.Личинки, що живуть у
грунті в місцях скупчення рослинних детритів (гнізда мишей, ховрашків,
скупчення перегною, сміття, стружок тощо), що активно розшукують корм і
годуються мертвими рослинними залишками. Сюди відносяться частково носороги
(Oryctes), кукурудзяні гнойовики (Pentodon), деякі бронзівки (Epicometis hirta,
Oxythyreafunesta, Potosia (Netocia) hungarica та інші види підроду Netocia,
деякі Aphodiini (Pleurophorm, Mothon, Aphodius plagiatas). Усі види цієї групи
в личинковій фазі нешкідливі, але багато з них шкодять у імагінальній фазі
3.Личинки, що живуть у
грунті та харчуються підземними грибами. Сюди належить Odontaeus armiger. В
усіх фазах нешкідливий.
4.Личинки, що живуть у
мурашниках та харчуються матеріалами мурашиного гнізда. Сюди належить Potosia
metallica. Личинка нешкідлива, жук пошкоджує квіти рослин.
5.Личинки, що живуть у
порохнявій деревині і деревній потерті та харчуються рослинними детритами. Сюди
відносяться всі Lucanidae, Dynastini (частково), Trichiini, Valgini, Cehniini.
Личинки нешкідливі, але деякі з них є проміжними хазяїнами паразитів домашніх
тварин, деякі види шкодять в імагінальній фазі.
6.Личинки, що живуть у
трупах тварин. Сюди відносяться види Trох. Нешкідливі.
7.Личинки, що розвиваються
в природних купах калу тварин і харчуються ним. Здатні до пересування в межах
даної купи. Сюди належить більшість личинок Aphodiini. Нешкідливі. Деякі види в
імагінальній фазі є проміжними хазяїнами паразитів людини.
8. Личинки, що живуть у
субстраті, підготовленому для них
батьками та харчуються ним і не здатні до відшукування собі їжі.
Сюди відносяться дві підгрупи:
а) ті, що живляться калом
тварин, заготовленим у виді ковбасок, груш тощо - гнойовики з триб Сорrіnі і
Geotrupini. Для культурних рослин нешкідливі. Деякі види в імагінальній фазі є
проміжними хазяїнами паразитів людини і домашніх тварин;
б) ті, що живляться
рослинною масою, заготовленою в грунті в особливих осередках - в імагінальній
фазі.
Тривалість генерації в
різних представників розглянутих родин коливається від двох до шести генерацій
на рік. Тут можна розрізнити такі типи:
1. Багаторічна генерація,
остання зимівля в імагінальній фазі. Наприклад: травневий хрущ (Melolontha) має
чотирьох - або п'ятирічну генерацію. Після перезимівлі літає весною і відкладає
яйця, з яких у те ж літо виходять личинки. Вони зимують тричі (при чотирирічній
генерації) або чотири рази (при п'ятирічній генерації), наприкінці літа
перетворюються на ляльок, восени виходить жук, що зимує. Квітневий хрущ
(Rhizotrogus aequinoctialis) та інші види цього роду мають трирічну генерацію.
Після перезимівлі жук літає весною і відкладає яйця. Личинки, які з'явилися в
те ж літо, зимують двічі, наприкінці літа перетворюються на ляльок, восени
виходять жуки, що зимують. Вірогідно, такий же тип генерації мають хрущі роду
Chioneosoma.
2. Багаторічна генерація,
усі зимівлі відбуваються у фазі личинки, яка після останньої перезимівлі
перетворюється на ляльок навесні; виліт жуків відбувається наприкінці весни або
влітку. Личинки з яєць відроджуються в те ж літо, коли були відкладені яйця.
Наприклад: жук-олень (Lucanus cervus) має шестирічну генерацію, личинка зимує 6
разів; жук-носоріг (Oryctes nasiicornis) має чотирирічну генерацію, личинка
зимує 4 рази; мармуровий хрущ (Polyphylla fullo) і волосатий хрущ (Anoxia
pilosa) мають трирічну, а іноді чотирирічну генерацію, личинки зимують 3 або 4
рази; червневий хрущ (Amphimallon solstitialis), хрущ Нордмана (Monotropus
nordmanni), хлібний кузька (Anisoplia austriaca) мають дворічну генерацію,
личинки зимують 2 рази.
. Однолітня генерація,
зимівля тільки в імагінальній фазі. Наприклад: оленка волохата (Epicometis
hirta), угорська бронзівка (Potosia hungarica), гнойовики з триб Соргіпі,
Geotrupini, кравчики (Lethrus), деякі Aphodiini.
. Однолітня генерація,
зимівля у фазі личинки. Наприклад: садовий хрущик (Phyllopertha horticola),
квіткоїди (Anomala), піскорийки (Норііа), листові хрущики (Homaloplia), багато
видів кузьок (Anisoplia segetum, A. leucaspis, A. deserticola), деякі бронзівки
(Cetonia aurata, Potosia metaliica), пістряки (Trichius, Gnorimus), кравчик
(Lethrus apterus), деякі Aphodiini.
.Однолітня генерація,
зимівля в личинковій та імагінальній фазах. Наприклад: рудий нічний хрущик
(Serica brunnea), шовковистий нічний хрущик (Maladera holosericea), золотава
бронзівка (Cetonia aurata), короткокрилий пістряк (Valgus hemipterus).
6. У році дві генерації.
Наприклад: деякі види Aphodius (Aphodius melanostictus, A. hydrochoeris).
Незалежно від тривалості
генерації усі личинки пластинчастовусих і близьких до них родин рогачів і
троксів мають 3 віки, тобто линяють тричі і після останнього линяння
перетворюються на ляльок. Личинки, які вийшли з яйця і ще не перелиняли,
називаються личинками 1-го віку, що перелиняли 1 раз - личинками 2-го віку і
перелиняли двічі - личинками 3-го віку. Розмір личинок поступово збільшується в
проміжках між линьками, причому їхні зовнішні покриви, крім сильно
хітинізованих частин (голови, вусиків, кінцівок), поступово розтягуються. Тому
хоча за розміром личинки і можна судити приблизно про її вік, але при цьому
можливі помилки, тому що розмір личинок уже після виходу з яйця може
розрізнятися, а личинка, яка щойно перелиняла, за розміром може не відрізнятися
від личинки попереднього віку перед линянням. При вимірі личинки проміряють її
довжину по спині від переднього краю лоба (основи верхніх щелеп) до анального
отвору. Більш надійне визначення віку личинки за розміром головної капсули - її
довжиною і шириною. Довжина виміряється від потилиці до переднього краю лоба
(без лиштви), ширина в найширшому місці голови - відразу за основою вусиків,
або по-серединта довжини, залежно від форми голови.
Якщо личинка перебуває в
одному віці понад рік, то визначити, скільки часу вона прожила, неможливо. За
станом верхніх щелеп можна лише довідатися, чи давно перелиняла дана личинка. У
личинки травневого хруща верхня щелепа неперелинялої або, навпаки, давно
перелинялої личинки відрізняється притупленим кінцем і наявністю западини, що
перетинає її верхню частину. Верхня щелепа недавно перелинялої личинки
відрізняється насамперед загостреним кінцем, а, крім того, верхня її частина на
деякій відстані від різального краю, перетинається двома поперечними
післялиньковими борозенками, від яких до різального краю мандибули тягнуться
численні післялинькові зморщечки. З часом унаслідок постійного тертя об грунт
кінець щелепи притупляється, післялинькові борозенки і зморщечки стираються і
щелепа стає такою, як у личинки, що не перелиняла
У ході досліджень
установлено, що в ШНПП можна зустріти личинок наступних видів пластинчастовусих
жуків: Amphimalon solstitialis, Cetonia aurata L., Geotrupes stercorarius L.,
Lethrus apterus Laxm., Melolonta melolonta L., Miltotrogus aequinoctialis
Hrbst, Miltotrogus vermis Germ., Onthophagus sp., Rhizotrogus aestivus Ol.,
Serica brunnea L.
Найбільш поширеними видами є: Травневий
хрущ (Mellolonta hippocastani), Нехрущ червневий (Amphimallon solstitialis),
Хрущик садовий.
У розподілі цих тварин за
біотопами можна виявити певну закономірність. Личинки Miltotrogus
aequinoctialis Hrbst., Rhizotrogus aestivus і Amphimalon solstitialis надають
перевагу чорноземним грунтам. Тому зазначені види уникають піщаних ґрунтів
арени, заплавних місцеперебуваннь і частіше можуть бути зустрінуті в лісах.
Личинки Lethrus apterus і Onthophagus sp. є копрофагами. Тому тип ґрунту не
виступає як важливий фактор, що визначає їх просторовий розподіл. Однак частіше
ці види зустрічалися на плакорі.
Легкі ґрунти сухуватого
бору і субору на арені заселяються хрущем Serica brunnea. Хрущі Melolonta
melolonta досить часто зустрічаються в лісових біогеоценозах Шацького
національного природного парку. Біотопами, яким ций вид надає перевагу, є
помірно вологі заплавні стадії.
У болотистих ґрунтах
притерасся хрущі не зустрічаються. Це пов'язано з тим, що личинки
пластинчастовусих належать до групи власнеґрунтових тварин. Вони здатні глибоко
проникати в ґрунтову товщу і переносити умови досить сильної сухості. Але
надлишкова вологість ґрунту є чинником, що лімітує розподіл цих тварин.
Таким чином, личинки
пластинчастовусих виявляють досить високий ступінь приуроченості до певних
типів ґрунтів і біогеоценозів. Тому можна виділити індикаторні групи видів.
Індикаторами чорноземного
типу ґрунтоутворення є види Miltotrogus aequinoctialis Hrbst., Rhizotrogus
aestivus і Amphimalon solstitialis. Індикатором лугового процесу в
ґрунтоутворенні є Melolonta melolonta. Індикатором дерено-борових ґрунтів є
Serica brunnea.
Туруни (Carabidae)
Дуже поширена група комах. Покриви сильно
склеротизовані, містять чорний або бурий пігмент з групи меланінів, який
обумовлює їхнє забарвлення - чорне або темно-буре. Поверхневий шар покривів
може бути гладким, але частіше несе мікро- і макроскульптуру. Макроскульптура дуже
різноманітна, може складатись із горбиків, зерняток, зморшок.
Макроскульптура складається із сітки
багатокутних осередків, що покривають поверхню. Покриви майже завжди несуть
щетинки і дуже часто волоски.
Голова з добре видним зверху, іноді дуже
великими мандибулами. До складу черепа входить 3 великі склерити: епікраніум,
епістом, базикраніум. З боків розташовані складні очі. Простих очей на лобі
немає.
Груди Передньоспинка - дорзальні склерити
передньогрудей - бувають більш-менш серцеподібні, рідше розширені до основи.
Середньо- і задньогруди тісно
зрощені між собою. Ступінь розвитку задньогрудей пов’язаний з розвитком задніх
крил.
Черевце. Являє результат еволюції черевця
первинних комах, яке складається з 11 сегментів (уритів).
Відіграють велике значення в біоценозах.
Більшість з них хижаки, які поїдають молюсків, дощових червів. Вдень ховаються
під камінням пістилки, відлущеною корою, вночі полюють на комах.
Найбільш поширеними представниками є:
Скакун лісовий - жук бронзо-чорного
забарвлення, розміри 14 - 19 мм з білими перев’язками. Живе у піщаних сухих
лісах.
Краснотіл бронзовий - тіло зеленкувате,
бронзово -чорного кольору. Довжина 16 - 21 мм.
Турун садовий (Carabus hortensis). Довжина
23 мм. На чорних надкрилах по 3 ряди блискучих ямочок. Живе в лісах, садах.
Турун чорний (Carabus nigra).
Зустрічається в лісі під корою пеньків, під листям. Типовий хижак.
Мокриці
Один із підрядів
рівноногих ракоподібних, представники якого, покинувши вихідне і нормальне для
більшості видів свого класу водне середовище, тією чи іншою мірою
пристосувались до наземного способу життя, причому деякі з них навіть до
екстремальних за режимом зволоження умов пустелі. Зокрема під наметом лісу їх
просторове розміщення визначається перш за все вологістю ґрунту та підстилки.
Ротові органи мокриць
гризучого типу, пристосовані до споживання цупкого грубого субстрату шляхом
відривання чи зіскоблювання та наступного подрібнення перед проковтуванням. Це
дозволяє їм повною мірою використовувати для їжі листовий опад, а деяким видам
навіть гнилу деревину.
На відміну від більшості
ґрунтових сапрофагів, мокриці малоперебірливі відносно ступеня попередньої
переробленості опаду, споживаючи з однаковою активністю як малозруйноване листя
з верхнього шару підстилки, так і з ферментативного, що вже зазнало істотного
впливу мікроорганізмів. Це пояснюють меншою їх чутливістю в порівнянні
наприклад з диплоподами та дощовими червами, до поліфенолів, що містяться у
свіжому опаді. Більше споживання корму може зумовлюватись не його перевагою, а
навпаки - браком в ньому поживних речовин взагалі, чи окремих незамінних
компонентів.
При харчуванні рослинними
рештками з низьким вмістом кальцію його брак компенсується поїданням тваринних
залишків, зокрема власних, скинутих при линці покривів і навіть канібалізмом.
Щодо міді, то її недостатність в рослинному кормі надолужується більшим його
споживанням, а також копрофагією. Коли трапляється нагода мокриці можуть
поїдати фекалії чи то власні, чи різних інших ґрунтових сапрофагів.
Мокриці являють зручний
приклад для аналізу актуальної для тварин -сапрофагів проблеми співвідношення у
живленні власне мертвих решток та живих мікроорганізмів, що їх насичують і
істотною мірою здійснюють розкладання. Збільшення кількості певних груп
мікроорганізмів та підвищення загальної їх функціональної активності в травному
тракті порівняльно до споживаного листового опаду свідчать, що мікроорганізми у
мокриць не тільки не перетравлюються, але навіть можуть тут розмножуватись.
Таким чином для мокриць головним джерелом живлення є не бактеріальна маса, а
мертві тканини споживаних макроорганізмів, переважно рослинних. Самі ж мікробні
клітини як було встановлено, перетравлюються мокрицями лише при тривалому
голодуванні, що є додатковим свідченням адаптивного характеру їх взаємин. Однак
слід зауважити, що хоча мікроорганізми в нормі не споживаються, і спеціально не
перетравлюються, вони все ж відіграють важливу роль у живленні мокриць як і
інших ґрунтових сапрофагів, для яких істотною складовою живлення є утворені під
дією мікробних ферментів продукти розкладання рослинного опаду.
.2 Хорологія грунтової
мезофауни у районі дослідження
3.2.1 характеристика
типових лісів Рожищенського району
Гетерогенність ґрунтового середовища необхідно
розглядати як фактор формування таксономічного й екологічного різноманіття
тваринного населення ґрунту. Важливу роль у формуванні тваринного населення
ґрунтового горизонту відіграє органічний матеріал, що надходить у вигляді
надземного опаду і маси відмерлого коріння. Тому, необхідно досліджувати
грунтову мезофауну у різних біогеоценозах.
На території Рожищенського району переважають
соснові ліси, які і нині становлять 77,1 % площі лісовкритих земель. Значну
частину резервату займають березові ліси - 16,8 %. Площа вкрита вільховими та
іншими лісами складає 6,1%.
У регіоні дослідження соснові ліси займають
дерново-слабопідзолисті піщані і супіщані грунти. Залежно від умов зволоження і
родючості грунтів всю різноманітність сосняків згідно класифікації Поварніцина
В. О. зводять до таких груп: лишайникової, зеленомошникової, довгомошникової,
сфагнової і складної.
Лишайникові сосняки, або сухі бори, займають
підвищені частини рельєфу і ростуть на сухих, порівняно малородючих
слабопідзолистих пісках з низьким рівнем підґрунтових вод.
Зеленомошникові, або свіжі і почасти вологі,
сосняки ростуть на середньозволожених слабопідзолистих піщаних або супіщаних
грунтах.
Довгомошникові, або вологі, сосняки пристосовані
до дуже зволожених дерново-підзолистих піщаних або супіщаних грунтів з слідами
оглеєння і високим рівнем ґрунтових вод.
Сфагнові сосняки, або сосна по болоту, дуже
поширені в північній чатистині заповідника. Вони займають знижені частини
рельєфу з торф'янисто-оглеєними грунтами, які підстилаються пісками.
Складні сосняки з добре розвиненим другим ярусом
з дуба, граба, липи і іноді підліском з ліщини ростуть на порівняно багатих
супіщаних, рідше піщаних грунтах з прошарками суглинку або з добре виявленим
суглинистим горизонтом
Досліджували безхребетних дерено-підзолистих ґрунтів,
що переважають в даних сосняках, і торфяно-болотних грунтах, типових для
регіону дослідження.
Розподіл дерено-підзолистих ґрунтів носить
зональний характер, вони займають 42,3%, відрізняються високою кислотністю і
вимагають посиленого вапнування. У природному стані по ступені виразності
підзолистого обрію ці ґрунти розділяються на слабко-, середньо- і
сильноопідзоленні.
Торф'яно-болотні ґрунти складають 15,02%, також
характеризуються високою кислотністю.
На суглинних і супіщаних різновидносях дерено-підзолистих
ґрунтів розміщаються головним чином багаторічні деревні насадження (91,6%). Сосняк
лишайниковий - Pineum cladinosum. Приурочений до підвищених територій.
Займає дерново-підзолисті слабо опідзолені та сильно кислі ґрунти. Ґрунти бідні
рухомими формами фосфору і калію, дещо більше кальцію. Сосна формує одноярусні
моно домінантні деревостої ІІІ - ІУ бонітету із запасом деревини в І класі віку
40-50 м3/га. Підлісок практично відсутній. Одно ярусність і невелика
повнота фітоценозу сприяють прониканню сонячних променів.
Сосняк лишайниковий зустрічається досить рідко,
займає верхні частини дюн, де тягнеться звичайно вузькою смужкою на
дерново-слабопідзолистих пісках із слабо виявленим гумусовим горизонтом.
Будова грунту:
А0 - пухка підстилка з хвої і дрібних
гілочок сосни, 1 - 3 см.
А1 - сухий пісок темносірого кольору,
переплетений дрібними корінцями рослин, 3 - 5 см.
А2 - сухий пісок світлосірого кольору,
5 - 8 см.
В1 - сухий пісок коричневого кольору,
знизу забарвлення стає більш
світле, 30 - 41 см.
В2 - сухий пісок пальового кольору, 80
- 100 см.
С - з глибини 120 - 150 см пісок білого кольору з
пальовим відтінком.
Деревостан у віці 40 - 50 років, зрідка старіше,
досягає висоти 6-8 м, має нерівномірну зімкнутість крон до 0,5 - 0,6, за
продуктивністю належить до IY/Y класу бонітету. Підлісок звичайно не виявлений,
інколи складається з поодиноко розкиданих кущів рокитника (Суtіsus Zingeriі V.
Кгесг.) і лем-ботропіса (Lembotropis nigricans).
Трав'яний покрив рідкий, ступінь вкриття 0,2 -
0,3; складається з типцю, чебрецю, до яких домішуються поодинокі екземпляри
золотушника, мучниці, нечуй-вітру, келерії, булавоносця, цмину, агалик-трави,
вересу, щавлю, гвоздики, мітлиці, смовді, лещиці.
Лишайниковий покрив добре виявлений, з ступенем
вкриття 0,8, інколи менше. Він складається із звичайних видів лишайників:
Сlаdопіа silvatica, Сl. rangiferina, до яких домішані: Сlаdопіа silvatica, Сl.
Ипсіаlis, Сl. fimbriata. Крім лишайників, звичайно у вигляді невеликих груп тут
зустрічаються зелені мохи: Discranum scoparium, D. Undulatum.
Відновлення під пологом насаджень тут проходить
погано. У підрості зустрічаються поодинокі екземпляри сосни віком 5 - 10 років,
заввишки 0,5 - 1,0 м.
Сосняк сфагновий: (Pineum sphagnosum) розміщений в
основному на верхових болотах. Нагромаджує і зберігає вологу. Вік 38 років,
бонітет II, повнота 0,71, запас 120, приріст 3,14 м3/га. Ґрунти тут
д торфяно-болотні із сильно застояними водами. У таких умовах виростає тільки
сосна.
Сфагнові сосняки дуже поширені у регіоні
дослідження, де вони займають значні площі в знижених частинах, рельєфу з
торф'янисто-оглеєними грунтами, які підстилаються пісками.
Утворення вказаних сосняків і їх продуктивність
залежить від товщини шару торфу, яка варіює від 15 до 100 - 150 см і більше.
Різниця між сфагновим сосняком і сфагновим
болотом полягає в тому, що сосняку властивий порівняно добре зімкнутий
деревостан, дерева якого впливають одно на одного. Коли ж деревостан сосни під
впливом великого шару торфу настільки розріджується, що поодинокі дерева в
надземній та підземній частинах не впливають одно на одного, тоді сфагновий
сосняк переходить в сфагнове болото.
Сфагнові сосняки займає знижені частини рельєфу з
торф'янисто-оглеєним грунтом, який підстилається піском. Товщина шару торфу тут
варіює від 15 до 70 - 100 см.
Деревний ярус складається з сосни, яка у віці 60
років має діаметр в середньому 12 см і заввишки 15 м. За продуктивністю
деревостан найчастіше належить до III класу бонітету, іноді при початкових
стадіях заболочування піднімається до II класу, а при дуже розвиненому шарі
торфу (близько 70-100 см) знижується до IV класу бонітету. Підлісок звичайно
відсутній, в трав'яно-чагарниковому покриві панують суцільні зарості багна, до
якого домішуються пухівка (Eriophorum uaginatum), журавлина, лохина, чорниця,
андромеда і а ін. Моховий килим складається із сфагнових мохів Sphagnum
recurvum з домішкою Sphagnum acutifolium та Sphagnum magellanicum.
Сосняк чорничний: (Pineum myrtillosum). Для
досліджуваної ділянки характерний знижений рельєф, піщані грунти (із вологістю
29,9%) з легким торф'янистим шаром. Склад деревостою І - го ярусу 9 - 10 С, 0 -
1 Б, висота 20 - 23 м, вік 48 - 50 років. Насадження високопродуктивним, ІІ
бонітетом. Зустрічається береза, висотою 1,5 - 2 м. В підліску поодинокі
горобина, верба, трав’янистий покрив - чорниця (90% покриття), місцями багно,
брусниня, мохи.
Приурочений до дещо зниженого рельєфу. Ґрунтова
різниця представлена дрібнозернистими пісками (найчастіше з невеликими
прошарками супісків) і характеризується високою гідролітичною кислотністю у
всіх шарах. Верхній обрій іноді відірваний. Ґрунтові води розташовуються на
глибині понад 1,4 м. Ґрунти більш вологі, чим в моховидному типі лісу,
сильнокислі у всіх обріях, багаті кальцієм, магнієм, калієм, фосфором,
нітратним і аміачним азотом, особливо у верхньому обрії, забрудненому
промисловими викидами, де відзначаються високі суми поглинених основ (17,50 -
20,60 мг-екв/100 м ґрунту).
Для ділянки характерні одноярусний древостій II
бонітету з запасом деревини в IV класі віку в середньому 200 - 300 м3/га
(повнота 0,6 - 0,7). Підлісок розвинутий задовільно, представлений вербою
сіркою, горобиною. У підростку одиноко зустрічаються ялина, береза. Древостій
75-80 років, висота його 21 м, середній діаметр 22-24м.
Сосняк зеленомошковий: (Pineum pleuroziosum) Місцевість дещо
підвищена, проте без горбкуватостей. Піщані грунти (вологість 6,9%).
Будова грунту:
А1 - пісок чорного кольору,
переплетений дрібними корінцями рослин, 10-15 см.
А2 - пухкий свіжий пісок світлосірого
кольору, 8 - 15 см.
В - свіжий пісок світло-коричневого кольору,
знизу забарвлення стає більш світле, 20 - 30 см.
С - з глибини 60-100 см пісок білого кольору.
Сполука древостою 10 С, висота 22 - 23 м, вік 45
- 48 років. Полог зімкнутий. Підріст не виражений, поодиноко зустрічається
сосна, береза, осика висотою до 1 м. Підлісок відсутній. В трав'яному покриві
переважають мохи (мох Шребера), займаючі до 96%, іноді зустрічається вереск,
брусниця, плаун. Для даного типу сосняку типовий слабий розвиток трав'янистого
покриву та бідність флористичного складу. В ньому у вигляді окремо розкиданих
екземплярів зустрічаються верес, біловус, брусниця, ожика, орляк, перестріч
лучний та інші рослини. Суцільний моховий покрив товщиною 3 - 5 см.
Характерною рисою підстилки соснових лісів
природних фітоценозів є зосередженість основних запасів чисельності і біомаси
безхребетних (до 85 %) у підстилці Ао і верхніх горизонтах А1.
Уже на глибині 20 - 25 см, а у сірих лісових - з 10-15 см щільність тварин
різко знижується до 8 - 24 екз/м2 і 1 - 4 г/м2.
У Рожищенському районі ми досліжували
грунтову мезофауну у соснових лісах лишайникових, сфагнових, чорничних,
мохових; березових лісах.
3.2.2 Поширення грунтової мезофауни у соснових
лісах
Структура ґрунтової мезофауни у значній мірі
залежить від стану фітоценозу, що під впливом різних факторів, і насамперед
абіотичних, зазанає змін.
Великий вклад в дослідження грунтових
безхребетних лісових біогеоценозів внесли роботи М.С. Гілярова, Ю.І. Чернова,
О.І. Криволуцького та ін. В цих роботах подається характеристика зонального
розподілу тваринного населення грунтів. При цьому наводяться параметри їх
чисельності, зоомаси і метаболізму. Однак грунтова фауна лісових біогоценозів
окремих регіонів країни все ж недостатньо вивчена. Це і послужило причиної
системного дослідження основних груп грунтової мезофауни Рожищенського
району .
Стабільність структури комплексу грунтової
мезофауни обумовлена широким спектром груп тварин, оптимальним співвідношенням їх
у еколого-трофічному ланцюгу. Нами проведено облік усіх наявних матеріалів
чисельності, біомаси і групового складу мезофауни.
М.С. Гіляров засвідчує, що за період другої
половини ХХ століття чисельність і біомаса педобіонтів змінилася незначно і
складає відповідно 150 - 400 екз/м2 і 32 - 67 г/м2 у
залежності від ґрунтового таксону. Зберігається високою насиченість підстилок
соснових лісів групами Lumbricidae, Chilopoda, Diplopoda, Aranea, Carabidae,
Elateridae, Curculionidae і набагато нижче - Tenebrionidae, Alleculidae,
Cicadidae і ін. З артропод найбільш поширені Carabus (L), Notiophilus Dum.,
Pterostichus Bon., Harpalus Latr., Philonthus Curt., Lathrobium Grav.,
Dorcadion Dalm. [9].
Фауністичний склад грунтової мезофауни у соснових
лісах регіону дослідження включає представників із 5 класів, 8 рядів, 14 родин,
42 видів. Тут менше всього дощових червів 4,4 екз/м2, як правило
вони є індикаторами типу сосняку де їх чисельність варіює від 0,8 до 7,6 % від
чисельності всіх безхребетних. Відсоток маси червів від загальної маси
безхребетних в сосняках чорничних, мохових та лишайникових відповідно
21,7-48,3; 6,1; 11,3-13,1.
Чисельність двопарноногих багатоніжок майже
однакова у всіх типах сосняків. Молюски в більшості гігрофіли і частіше
зустрічаються у вологих сосняках, зокрема чорничних - 5,7 екз/м2.
Чисельність павукоподібних досягає 10 - 50 екз/м2
в окремих біогеоценозах вони домінують.
Серед комах практично у всіх типах сосняків
переважають твердокрилі, жужелиці та стафіленіди. Ковалики зустрічаються у всіх
ґрунтах сосняків, представлені 7 видами, серед яких, домінуючими видами є
Athous subfuscus та Dalopius marginatus. Вони домінують в чорничних і мохових
сосняках і досягають 58% від числа всіх твердокрилих (рис. 2.). В сосняках
сфагнових та лишайникових їх чисельність невелика і складає біля 5%.
Довгоносики живуть у порівняно сухих лишайникових
сосняках, де їхня чисельність досягає 30 екз/м2. Вони представлені 3
видами. Основну масу їхніх личинок складає Sirophosoma capitatum. У більш
вологих чорничних сосняках чисельність довгоносиків невелика.
Чисельність коротконадкрилих жуків у всіх типах
сосняків досягає 6,1-19,6 екз/м2. Видові угрупування їх більш
різноманітна, чим інших родин твердокрилих, але в порівнянні з фауною листяних
лісів трохи збіднений. Стафіленіди нараховують 12 видів: з них 6 домінують,
однак чисельність їх не перевищує 4,8 екз/м2. Найбільш численний
Drusilla canaliculata.
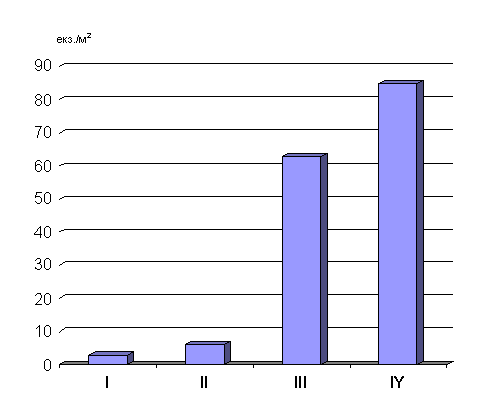
І - сосняк лишайниковий; ІІ - сосняк сфагновий;
ІІІ - сосняк чорничний; ІY - сосняк моховий
Рис. 2. Чисельність Elateridae в різних соснових
лісах
Чисельність жужелиць близько 14 екз/м2.
В усіх типах сосняків вони складають у середньому 9,6% чисельності твердокрилих
(у сфагнових - 14,9%, у чорничних - 8,7, у моховидних і лишайникових - 7,2 і
7,8 %). Жужелиці представлені 9 видами. По чисельності виділяються Calathus
micropterus, Calathus arvensis, Epuphius secalis, Agonum obscurum. Чисельність
інших родин жуків (пластинчатовусі, м’якотілки та листоїди) у всіх типах
сосняків невелика, це стосується і інших безхребетни, що ілюструє рисунок 3.
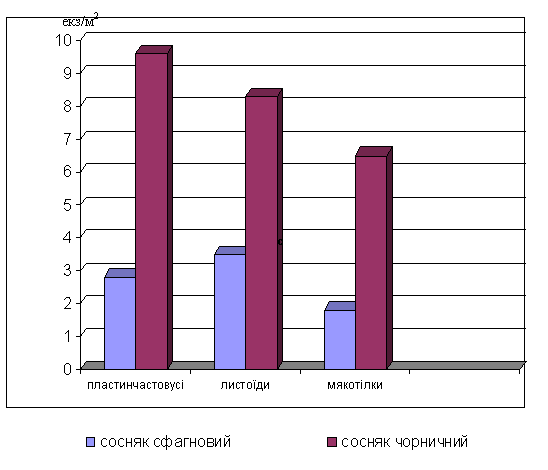
Рис. 3. Чисельність пластинчастовусих, м’якотілок
та листоїдів в соснових лісах
Довгоніжки були виявлені тільки в вологих типах
сосняків. Товстоніжки нечисельнні в соснових лісах. В чорничних типах сосняків порівняно часто зустрічаються
бекасниці (у більш сухих типах сосняків 22 -31 екз/м2). Справжні
мухи в незначній кількості знайдені тільки в сосняках лишайникових.
Біомаса ґрунтової мезофауни в цілому в сосняках
різних типів не більша 2,6 г/м2. У комах вона варіює в межах 29,9 -
85,0%, а у дощових червів - 11,3 - 25,6% (в окремих типах сосняків).
Виділяються по зоомасі також твердокрилі (72,3 - 88,5%) і двокрилі (3,1 -
12,4%) (рис. 5.2).
Сама висока біомаса в різних типах сосняків у
коваликів - 0,73 г/м2. Зоомаса жужелиць порівняно невелика, хоча в
ряді сосняків чорничних досягає приблизно 0,45 г/м2. Зоомаса
стафілинід ще менша - 0,34 г/м2.
Біомаса довгоносиків в менш вологих сосняках
іноді сягає 0,3 г/м2 .
Біомаса павуків максимальна в сосняках чорничних
0,39 г/м2 і мінімальна і сосняках зеленомошкових і лишайникових 0,20
г/м2. До деякої міри павуки є індикаторами вологості ґрунту і підстилки.
Біомаса губоногих і двопарноногих багатоніжок
міняється відповідно до їхньої чисельності в відповідності із чисельністю і в
різних типах сосняків практично однакова. Лише в чорничних сосняках зростає до
0,52 г/м2. Інші групи комах дуже нечисленні в сосняках.
Видова різноманітність мезофауни найвища в більш
вологих сосняках сфагнових та чорничних (3,60) і найнижчі в більш ксерофільних
типах сосняків лишайникових (1,89).
Показник вирівненості особливо високий у сосняках
сфагнових. Це свідчить про порівняно рівномірний кількісний розподіл видів
ґрунтових безхребетних.
Ми проаналізували наявність видів різних
зоогеографічних елементів грунтової мезофауни в сосняках регіону дослідження,
що вісвітлено у таблиці 1. Встановили, що серед них в сосняках переважають види
з більш вузькими ареалами.
Встановлено 7 зоогеографічних елементів:
європейський (35,1%), транспалеарктичний (30,8%), європейсько-сибірський (21,3%),
голарктичний, космополітний і европейсько-середземноморський (у цілому 12,8%).
Домінуючими є види європейські, транспалеарктичні та європейсько-сибірські, що
відображено у рисунку 4.
Більшість видів ґрунтових безхребетних сосняків
належить до лісової екологічної групи (84,2%); по 1-7% припадає на
лучно-лісову, лучно-польову, еврібіонтну, польову, болотну, лучно-болотну і
лучну групи.
Таблиця 1.
Види різних зоогеографічних елементів грунтової
мезофауни в сосняках району дослідження, %
|
Зоогеографічний
елемент
|
Сосняк
|
|
Сфагновий
|
Чорничний
|
Моховий
|
|
Космополітний
|
-
|
1,3
|
3,3
|
|
Голарктичний
|
9,7
|
8,7
|
8,3
|
|
Транспалеарктичний
|
29,2
|
35,0
|
28,3
|
|
Европейсько-арктичний
|
-
|
1,3
|
-
|
|
Европейсько-сибірський
|
27,8
|
16,2
|
20,0
|
|
Европейсько-середземноморський
|
1,4
|
2,5
|
1,7
|
|
Европейський
|
31,9
|
35,0
|
38,4
|
У сосняках по чисельності і біомасі переважають
зоофаги (72,5%), набагато менше фітофагів (13,7%) і детритофагів (9,9%), зовсім
мало копрофагів (3,8%), що відображено в рисунку 5. Бідність сосняків
сапрофагами можна пояснити відсутністю в підстилці листяного опаду, незначним
змістом у ґрунті гумусу і незначною придатністю соснової хвої для живлення
ґрунтових тварин.

Рис. 4. Види різних зоогеографічних елементів грунтової
мезофауни в соснових лісах, %

Рис. 5. Співвідношення чисельності трофічних груп
грунтової мезофауни в соснових лісах, %
.2.3 Поширення
грунтової мезофауни у березових лісах
Фауністичний склад грунтової мезофауни у
березових лісах Рожищенського району включає представників із 5 класів, 11
рядів, 17 родин, 64 видів. Середня чисельність грунтової мезофауни в
березняках, утворених березою бородавчатою, досить висока і складає 244,0 -
253,7 екз/м2.
Дощові черви та комахи займають по чисельності
домінуюче становище в березових лісах. Серед комах преважають твердокрилі і
двокрилі. Перші представлені 8 родинами; 4 із них домінують (довгоносики,
жужелиці, ковалики, стафілініди).
Найбільш багаточисельною групою твердокрилих в
березняках є личинки і жуки довгоносиків. В березняках мохових їх чисельність
сягає 98 екз/м2. Серед них домінують личинки щетинистого гостроокого
слоника, Otiorhynchus repleus, Phylloobius pyri. Чисельність жужелиць в
березняках різнив типів однакова (7,1 - 8,9 екз/м2), що не можна
сказати про стафілінід.
Ковалики в грунтах березняків представлені 10
видами. Їх чисельність в різних типах березняків варіює від (9,1 - 21,5 екз/м2).
Домінують серед коваликів рижуватий і обрамлений. Чисельність видів інших родин
жуків (пластинчастовусі, мікотілки, листоїди) незначна. Домінуючий в березняках
ряд двокрилих представлений 6 родинами.
Серед них переважають довгоніжки, бекасниці,
волочниці та псевдоволочниці. Довгоніжки надають перевагу більш вологим
біотопам, де їх чисельність складає 2,8 екз/м2. Чисельність
бекасниць в різних типах березняків коливається в межах від 2,0 до 6,1 екз/м2,
волочниць - приблизно однакова (2,9 - 5,1 екз/м2). Найменше
псевдоволочниць (2,7 екз/м2). Вони виявлені лише в березняках
чорничних, де переважають над іншими родинами двокрилих. Дещо менше в різних
типах березняків павуків і двопарноногих багатоніжок. Дуже малочисельні в
березняках пильщики.
Зазначимо, що в березняках переважають транспалеарктичні
(38,6) і європейські (53,8%) види, що продемонстровано у рисунку 6.

Рис. 6. Види різних зоогеографічних елементів грунтової
мезофауни в березових лісах району дослідження, %

Рис. 7. Співвідношення чисельності трофічних груп
грунтової мезофауни в березових лісах, %
При оцінці чисельності грунтової мезофауни, які
належать по певної трофічної групи, виявилося, що кількість фітофагів в
березняках чорничних становить 18,6% значно нижча, ніж в сосняках мохових,
сапрофагів - 23,4%, а зоофагів - 53,5, некрофаги - 4,5%, що продемонстровано у
рисунку 5.7.
ВИСНОВКИ
. Встановлений фауністичний склад представників
ґрунтової мезофауни у домінуючих лісах Рожищенського району. Всього виявлено
110 видів ґрунтової мезофауни, які належать до наступних таксономічних груп:
Комахи (Insecta), Багатоніжки (Myriapoda), Олігохети (Oligochaeta), Молюски
(Mollusca), Ракоподібні (Crustacea), Павукоподібні (Araneamorpha).
. Проаналізовано екологічні особливості поширення
грунтової мезофауни у різних типах лісу Рожищенського району, зокрема, у
сосняках лишайникових (Pineum cladinosum), сосняках сфагнових (Pineum sphagnosum),
сосняках чорничних (Pineum myrtillosum), сосняках мохових (Pineum
pleuroziosum), а також березових лісах.
. Встановлено кількісний склад і біомасу
представників ґрунтової мезофауни у соснових та березових лісах Рожищенського
району.
. Фауністичний склад грунтової мезофауни у
соснових лісах Рожищенського району включає представників із 5 класів, 8 рядів,
14 родин, 42 види.
Домінуючими представниками ґрунтової мезофауни
соснових лісів є комахи, серед яких практично у всіх типах сосняків переважають
твердокрилі, жужелиці та стафіленіди. Малочисельними є олігохети - 4,4 екз/м2,
які слугують індикаторами соснових типів лісів, де їх чисельність варіює від
0,8 до 5,6 % від чисельності всіх безхребетних.
Досліджено, що найменше представленою є ґрунтова
мезофауна у сосняках лишайникових (Pineum cladinosum).
. Найкраще розвинутою є ґрунтова мезофауна у
березових лісах. Фауністичний склад грунтової мезофауни у березових лісах
Рожищенського району включає представників із 5 класів, 11 рядів, 17 родин, 64
видів.
Середня
чисельність грунтової мезофауни в березняках, досить висока і складає
244,0-253,7 екз/м2.
Дощові черви та комахи займають по чисельності
домінуюче становище в березових лісах. Серед комах преважають твердокрилі і
двокрилі. Перші представлені 8 родинами; 4 із них домінують (довгоносики,
жужелиці, ковалики, стафілініди).
4.При оцінці чисельності грунтової мезофауни, яка
належать по певної трофічної групи, виявилося, що кількість фітофагів в
березових лісах становить 18,6%, сапрофагів - 23,4%, а зоофагів - 53,5,
найнижчі величини мають некрофаги - 4,5%. У соснових лісах по чисельності і
зоомасі переважають зоофаги (72,5%), набагато менше фітофагів (13,7%) і
детритофагів (9,9%), незначна чисельність копрофагів (3,8%). Бідність сосняків
сапрофагами можна пояснити відсутністю в підстилці листяного опаду, незначним
змістом у ґрунті гумусу і незначною придатністю соснової хвої для живлення
ґрунтових тварин.
5.Проаналізували наявність видів різних
зоогеографічних елементів грунтової мезофауни в соснових лісах регіону
дослідження. Встановлено 7 зоогеографічних елементів: європейський (35,1%),
транспалеарктичний (30,8%), європейсько-сибірський (21,3%), голарктичний,
космополітний і европейсько-середземноморський (у цілому 12,8%). Домінуючими є
види європейські, транспалеарктичні та європейсько-сибірські. Встановили, що
серед них в сосняках переважають види з більш вузькими ареалами. В березових
лісах переважають транспалеарктичні (38,6) і європейські (53,8%) види.
СПИСОК ВИКОРИСТАНОЇ ЛІТЕРАТУРИ
1. Алимов А.Ф. Молюски и их роль в биоценозах и формировании
фаун [Текст] : монографія / А.Ф. Алимов. - Л. : Наука, 1981.
- 248 с.
2. Андрусов Л. Материалы для фауны Oligochaeta окрестностей г.
Киева / Л. Андрусов // Зап. Киевск. об-ва естествоиспыт. - Киев, 1914. - Т. 23.
- Вып. 4. - С. 91-95.
3. Анисимова О.А. Хвойные деревья и насекомые-дендрофаги [Текст] : монографія /
О.А. Анисимова. - Иркутск, 1978. - С. 62-67.
4. Арзамасов И. Т. Влияние мелиорации на животный мир
Белорусского Полесья [Текст] : монографія / И.Т. Арзамасов, М.С.
Долбик, Э.И. Хотько. - Минск, 1980. - 369 с.
5. Бусленко Л.В. Життєдіяльність люмбріцід (Lumbrisidae
Oligochaeta Annelida) в дефльваних дерново-підзолистих грунтах Західного
Полісся / Л.В. Бусленко // Науковий вісник Волинського держ. ун-ту ім. Лесі
Українки. - Луцьк, 2008. - № 5 : Біологічні науки. - С. 46-50.
6. Веремеев В.Н. Видовой состав и численность почвенной
мезофауны сосновых молодняков Белорусского Полесья [Текст] : монографія / В.Н.
Веремеев. - Москва, 1980. - 515 с.
7. Гаврилюк В.С. Фізико-географічне районування Українського
Полісся [Текст] : монографія / В.С. Гаврилюк. - К.: Вид-во
Київ.ун-ту, 1955. - С. 59-65.
8. Гиляров М.С. Роль почвенных животных в формировании
гумусного слоя почвы / М.С. Гиляров // Усп. совр. биол. - 1951. - 31. - Вып. 2.
- С. 161-169.
9. Гиляров М.С. Животные и почвообразование / М.С. Гиляров
// Биология почв Северной Европы. - М. : Наука, 1988. - С. 7-16.
. Гиляров М.С. Зоологический метод диагностики почв [Текст]
: монографія / М.С. Гиляров. - Москва, 1965. - 265 с.
. Долгин В.Г. Определитель личинок жуков-щелкунов фауны
СССР [Текст] : монографія / В.Г. Долгин. - К., 1978. - 380 с.
. Демченко Е.Н. Состояние мезофауны беспозвоночных в почвах
[Текст] : монографія / Е.Н. Демченко. - К., 1984. - 125 с.
13. Іванців В.В. Структурно-функціональна організація комплексів
грунтових олігохет західного регіону України [Текст] : монографія /
В.В. Іванців. - Луцьк: РВВ „Вежа”, 2007. - 400 с.
14. Зражевский А.И. Дождевые черви как фактор плодородия
лесных почв [Текст] : монографія / А.И. Зражевский. - К. : Изд-во АН УССР,
1957. - С. 271.
15. Літопис природи. Рожищенський район // [Коніщук В.В. та ін.]. - Рожище, 2003. - Т. 1.
- 163 с.
16. Кипенварлиц А.Ф. Изменение почвенной фауны низинных болот
под влиянием мелиорации и сельскогозяйственного освоения [Текст] : монографія /
А.Ф. Кипенварлиц. - Минск, 1961. - 267 с.
17. Козлов М.А., Олигер И.М. Атлас - определитель безпозвоночных
[Текст] / М.А. Козлов,
И.М. Олигер. - М. : Просвещение, 1991. - С. 13-29.
18. Кулагин Н.М. К фауне русских Oligochaeta / Н.М. Кулагин //
Труды Зоологического отделения об-ва любителей естествознания. - 1886. - № 1. -
С. 1-65.
19. Мазурмович Б.М. Розвиток зоології на Україні [Текст] :
монографія / Б.М. Мазурмович. - Київ, 1972. - С. 78-94.
. Малевич И.И. К фауне Оligohaeta Белорусской ССР / И.И.
Малевич // Сб. трудов гос зоологич музея МГУ. - 1937. - 342 с.
21. Малевич І.І. Дощовики (Lumbricidae) лісових насаджень
України / І. І. Малевич // Праці ін-ту лісівн. АН УРСР - Т.ІІІ. - 1952. - С. 145-164.
22. Перель Т.С. Зависимость численности и видового состава
дождевых червей от породного состава лесонасаждений / Т.С. Перель // Зоолог.
журнал. - 1958. - № 37. - Вып. 9. - С. 1307-1315.
23. Песенко Ю.А. Принципы и методы количественного анализа в
фаунистических исследованиях [Текст] : монографія / Ю.А. Песенко. - М., 1982 -
85 с.
24. Піндрус О.М. Біологічні процеси та чинники розкладання
листового опаду як основа методики його компостування в зеленому господарстві
міста / О.М. Піндрус, П.П. Яворівський, О.В. Лукаш. - К.: Інститут зоології
НАНУ, 2004. - 108 с.
25. Попов В.П. Физико-географическое районирование Украинской
ССР / В.П. Попов, А.М. Маринич, А.И. Ланько. - К. : Київ. ун-т, 1968. - 683 с.
26. Плавильщиков Н.Н. Определитель насекомых [Текст] :
монографія / Н. Н. Плавильщиков. - М., 1950. - 544 с.
. Пахомов О.Є. Функціональні розмаїття ґрунтової мезофауни
заплавних степових лісів в умовах штучно забрудненого середовища [Текст] : монографія / О.Є.
Пахомов, О.М. Кунах. - Дп.: Вид. - во Дніпропетр. нац. ун - ту, 2005. - 324 с.
. Смоляк Л.П. Болотные леса и их мелиорация [Текст] : монографія / Л.П.
Смоляк. - Минск, 1969. - 258 с.
. Стадиченко А.П. К фауне и экологии пресноводных молюсков
Украинского Полесья / А.П. Стадиченко // Гидробиологический журнал. - 1984. -
№2. - С. 36-40.
30. Старобогатов Я.И. Bithynionidae фауны СССР [Текст] : монографія / Я.И.
Старобогатов. - Л. : Наука, 1987. - 168 с.
. Старобогатов Я.И. Клас брюхоногие молюски Gastropoda /
Я.И. Старобогатов. - Львов, 1977. - С. 5-12.
32. Стриганова Б.Р. Питание почвенных сапрофагов [Текст] : монографія /
Б. Р. Стриганова. - М. : Наука, 1980. - 244 с.
33. Стриганова Б.Р. Комплексы почвообитающих беспозвоночных в
пойме среднего течения Днестра / Б.Р. Стриганова // Зоол. журнал. - 1968. - Т.
47. - Вып. 3. - С. 56-62.
34. Сукачев В.Н. Основы лесной биогеоценологии / В.Н. Сукачев.
- М., 1964. - С. 5-49.
. Фасулати К.К. Полевое изучение наземных беспозвоночных
[Текст] : монографія / К.К. Фасулати. - М. : Мир, 1975 - 305 с.
36. Чернов Ю.И. Природная зональность и животный мир суши [Текст] : монографія /
Ю.И. Чернов. - М. : Мысль, 1975 - 222 с.
37. Чекановская О.В. Дождевые черви и почвообразование [Текст] : монографія /
О.В. Чекановская. - М. - Л. : АН СССР, 1960. - 206 с.
38. Хотько Э.И. Почвенная фауна Беларуси [Текст] : монографія / Э.И. Хотько. - Минск, 1993.
- 253 с.
39. Эйтминавичюте И.С. Закономерности формирования комплексов
почвенных беспозвоночных под влиянием антропогенных воздействий в зоне
дерново-подзолистых почв [Текст] : автореф. дис… д-ра биол. наук. / И.С.
Эйтминавичюте. - М., 1982. - 40 с.
40. Шапошникова Е.В. Дождевые черви в биоценозах западных
областей УССР / Е.В. Шапошникова // Проблемы почвенной зоологии: Тез. докл. VІІ
Всесоюзного совещания. - К., 1981. - С. 253.
41. Царик И.В. Роль почвенных беспозвоночных в разложении
растительных остатков в некоторых экосистемах Украинских карпат / И.В. Царик. -
М. : Наука, 1975. - С. 73-75.
. Adamovicz Z. Materialy do fauny mieczakow (Mollusca). -
Polesia. Mus. Zool. Pol. - 1939. - 146 р.
43. Baker F. The molluscan family Planorbidae. Urbana: Univ.
Of Illinois press, 1945. - 530 p.
44. Baкowski J. Mieczaki.- Lwow: Mus.im.Dzieduszyckich, 1892. -
264 s.
45. Belke G. Catalogue des animaux qui habitent le districh de
Radomysl // Bulletin de la Societe Imperiale des Naturalistes de Moscou. -
1866. - № 3. - 46-61 s.
46. Eichwald E. Naturhistorische Skizze von Lithauen,
Volhynier und Podolien in geognostisch - mineralogischer botanischer und
zoologischer Hinsicht. - Wilna. Zawadzki, 1830. - 256 s.
47. Kasprzak K. Skaposzczety Glebowe, ІІІ, Rodzina: Dźdźownice
(Lumbricidae). - Warszawa, Państwowe
wudavnichctwo naucowe, 1986. - T. 6. - 187 s.
48. Lomniski M. Materialy do fauny szaranzakow galicyjskich //
Sprawozdanie Komisyi fizijograficznes. - Krakow. - T. IX. - 1875. - p.1-14.
49. Moszyńsri A., Moszyńsra M. Skaposzczety (Oligochaeta). - Prirodnik Lwowski. - Pr.
Komis. Biol. PTPN, 1957. - T. 18. - №6. - S. 1-204.
50. Michaelsen W. Zur Kenntnis der Lumbriciden und ihrer
Verbreitung // Ежегодник Зоолог. музея Импер. Академии наук. - СПб., 1910. - №
15. - С. 1-74.
. Plisko J.D. Lumbricidae. Dzdzownice Fauna Polski. -
I.P.W.N. Warszawa, 1973. - 155 S.
52. Nowicki M. O rybach dorzeczy Wisly, Styru, Dniestry i Prutu w
Galicyi. - Krakow, 1889. - 54 s.
53. Nusbaum J. Zur Anatomie und Systematik der Enchytraeiden //
Biol. Zentralbl. - 1895. - № 15. - S. 25-31.
54. Nusbaum J. Materiały do historyi
naturalnej skąposzczetów (Oligochaeta) galicyjskich // Spraw. Kom. fizjogr. PAU,
Kraków, 1896. - № 31. - S.
9-62.
. Christian E., Zicsi A. Ein synoptischer Bestimmungsschlüssel der Regenwürmer Ostterreichs
(Oligochaeta: Lumbricidae) // Bodenkultur. - 1999. - №2. - S. 121-131.
. Yaworowski A. Fauna stadzienna miast Krakówa i Lwowa. - Spraw. Kom. Fiz., Kraków, 28. - S. 29-48.
57. Westerlund. C.A. Fauna der in der Palaarktischen Region
Lebenden Binnenconchylien: V. Fam. Succinidae. Auriculidae. Limnaejdae.
Cyclosnomidae. Hydrocenidae. - Lund : Hakan Ohisson, 1885. - 135 S.
ДОДАТОК А

Рис. А. 1. Карта-схема Черемського природного
заповідника:+ місця відбору
проб
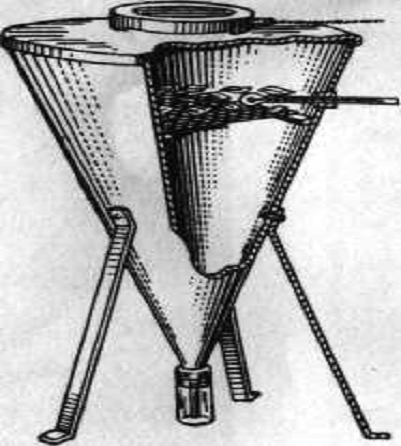
Рис. А. 2. Термоелектор в розрізі
(із Ріхтера)
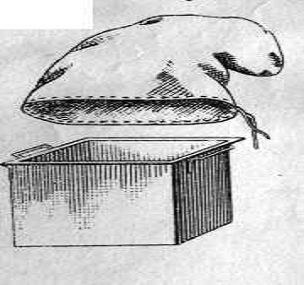
Рис. А. 3. Біоценометр Баскіной і
Фрідман
ДОПОВІДЬ 1
Шановний голово Державної екзаменаційної комісії,
шановні
члени Державної комісії, присутні. На Ваш розгляд
пропонується
дипломна робота на тему: „Мезофауна Черемського
природного заповідника”
Ґрунт - природне біокосне тіло, поряд з такими
його складовими, як тверда, рідка і газоподібна фази, містить і живу частину.
До складу ґрунтової біоти входять представники всіх царств живої природи.
Ґрунтове населення не тільки живе в ґрунті,
використовуючи його як середовище існування, але й активно трансформує його.
Тому цілком закономірне твердження, що ґрунт - це середовище існування й
одночасно продукт діяльності живих істот, що його населяють.
Ґрунтова біота - найбільш динамічна ланка
екосистеми, яка зазнає періодичних змін і, у той же час, що володіє певним
гомеостазом завдяки наявності механізмів регуляції і контролю.
Грунтові біоценози представлені
найрізноманітнішими безхребетними тваринами, а саме: найпростішими
(саркодовими, джгутиковими, інфузоріями), нематодами, коловертками, олігохетами
(люмбрицідами і енхитреїдами), черевоногими молюсками, хеліцеровими (павуками,
кліщами, косариками, псевдоскорпіонами), ракоподібними, багатоніжками,
комахами. При урахуванні особливостей життя і впливу на грунт тварин різних
розмірів їх поділяють на три групи: мікрофауну, мезофауну та мегафауну.
Мезофауна є важливим компонентом біогеоценозів.
Вона займає ведуче місце в зоомасі в складі ценозу, істотно впливає на процеси
трансформації органічних решток, міграції хімічних елементів в горизонтах
ґрунтового профілю, ґрунтоутворення, стимулювання біологічних процесів та
сукцесії організмів в біогеоценозах ґрунту. Без обліку мезофауни населення
ґрунту неможливо скласти загальний кадастр тваринного світу. Разом з тим,
мезофауна ґрунту включає значну кількість лісових і сільськогосподарських
шкідників, а тому має велике практичне значення.
Серед представників мезофауни ґрунтових
безхребетних тварин є багато стенобіонтних видів. Власне вони відповідають
вимогам індикації стану оточуючого середовища і можуть служити об‘єктами
моніторингу.
Розв‘язати завдання в напрямку біоіндикаційних
цілей можна лише на основі дослідження угрупування ґрунтових безхребетних в
цілому, в тому числі, представників мезофауни.
Відсутність праць, присвячених угрупуванням безхребетних
ґрунтової мезофауни Черемського природного заповідника, спонукало нас до
виконання досліджень спрямованих на вивчення даної групи тварин.
Актуальність дослідження зумовлена недостатнім
рівнем відомостей про фауністичний склад, особливості екології та поширення
ґрунтової мезофауни Черемського природного заповідника.
Об’єктом дослідження є ґрунтова мезофауна
Черемського природного заповідника.
Предметом дослідження є фауна, екологія,
особливості поширення ґрунтової мезофауни Черемського природного заповідника.
Мета роботи: вивчити еколого-фауністичні
особливості ґрунтової мезофауни району дослідження.
В роботі ставились такі завдання:
. Проаналізувати літературні джерела та
ознайомитись з історією вивчення ґрунтової мезофауни.
. Розглянути фізико-географічні умови Черемського
природного заповідника.
. Встановити фауністичний склад ґрунтової
мезофауни Черемського природного заповідника.
. Вивчити екологію ґрунтової мезофауни у районі
дослідження.
. Виявити особливості хорології мезофауни у
типових біогеоценозах соснових та березових лісів Черемського природного
заповідника.
Апробація роботи проводилася при захисті курсових
робіт.
Наукова новизна дослідження. Вперше вивчено
фауну, екологію та особливості поширення грунтової мезофауни у Черемському природному
заповіднику.
Практичне значення дипломної роботи полягає в
тому, що його результати можуть бути використанні у професійній діяльності
педобіологів, ґрунтознавців, агрономів, працівників сільського господарства та
лісництва, а також у навчальному процесі. Дані роботи можна використовувати
також при проведенні інтродукції та акліматизації окремих видів у відповідності
до типів ґрунтів.
У першому розділі висвітлена історія дослідження
ґрунтової мезофауни в Україні та суміжних територій, зокрема, розглянуто
ступінь вивчення фауністичного складу ґрунтової мезофауни регіону дослідження,
особливості поширення та вплив екологічних факторів на життєдіяльність цієї
групи тварин.
У другому розділі охарактеризовано основні
складові частини природних фізико-географічних умов Черемського природного
заповідника, а саме: геологічна будова території, рельєф, клімат, ґрунти, флора
і фауна. Все вказує на своєрідний генезис території та складний процес
створення природних територіальних комплексів, що істотно відбивлося на
формуванні мезофауни.
Матеріал для дослідження був зібраний протягом
2006-2008 рр. у домінуючих лісах Черемського природного заповідника.
Збір ґрунтової мезофауни здійснювали за
стандартними педобіологічними методиками М.С. Гілярова та К.К. Фасулатті. Це
методики прямого обліку, що дозволяють визначити чисельність ґрунтових тварин у
всьому заселеному ними обсязі ґрунту (до глибини зустрічальності) на одиницю
площі поверхні, або на одиницю об'єму грунту. В районі дослідження, нами були
обстежені в даний період різні групи мезофауни. Вибір прийомів для обліку
різних груп мезофауни визначався особливостями ґрунту й об'єктами дослідження.
Зокрема, були застосовані метод грунтових
розкопок, ловчих ліній, а також спеціальні методи обліку дрібних членистоногих.
При ідентифікації ґрунтової мезофауни ми
використовували визначники Н.Н. Плавільщікова (комахи), Е. Крістіана, А. Зічі
(черви), А.П.Стадніченко, Н.В. Сверлової, Р.І. Гураль (молюски).
У розділі „ЕКОЛОГО-ФАУНІСТИЧНА ХАРАКТЕРИСТИКА
МЕЗОФАУНИ Черемського природного заповідника" подані відомості щодо
видового складу ґрунтової мезофауни регіону дослідження та особливостей
екології досліджуваних видів.
Встановлений фауністичний склад представників
ґрунтової мезофауни у домінуючих лісах Черемського природного заповідника.
Всього виявлено 106 видів ґрунтової мезофауни, які належать до наступних
таксономічних груп: Олігохети (Oligochaeta), Комахи (Insecta), Багатоніжки
(Myriapoda), Ракоподібні (Crustacea), Павукоподібні (Araneamorpha), Молюски
(Mollusca).
У розділі «
ХОРОЛОГІЯ МЕЗОФАУНИ У БІОГЕОЦЕНОЗАХ Черемського
природного заповідника» охарактеризовано основні типи лісових
біогеоценозів регіону дослідження. Встановлено, що на території резервату
переважають біогеоценози соснових лісів, які становлять 77,1 % площі лісовкритих
земель. Значну частину резервату займають березові ліси - 16,8 %. Площа вкрита
вільховими та іншими лісами складає 6,1%.
Тому ми досліжували екологічні особливості
поширення грунтової мезофауни у домінуючих типах лісів, а саме: у сосняках
лишайникових, сфагнових, чорничних, мохових; березових лісах.
Фауністичний склад грунтової мезофауни у соснових
лісах ПДПЗ включає представників із 5 класів, 8 рядів, 14 родин, 42 види. Тут
менше всього дощових червів 4,4 екз/м2, як правило вони є
індикаторами типу сосняку де їх чисельність варіює від 0,8 до 7,6 % від
чисельності всіх безхребетних.
Серед комах практично у всіх типах сосняків
переважають твердокрилі, жужелиці та стафіленіди.
Видова різноманітність мезофауни найвища в більш
вологих сосняках сфагнових та чорничних (3,60) і найнижчі в більш ксерофільних
типах сосняків лишайникових (1,89), зокрема це продемонстровано на прикладі
коваликів РИС. 1
Фауністичний склад грунтової мезофауни у
березових лісах ПДПЗ включає представників із 5 класів, 11 рядів, 17 родин, 64
видів.
Середня чисельність грунтової мезофауни в
березняках, утворених березою бородавчатою, досить висока і складає 244,0-253,7
екз/м2.
Дощові черви та комахи займають по чисельності
домінуюче становище в березових лісах. Серед комах преважають твердокрилі і
двокрилі. Перші представлені 8 родинами; 4 із них домінують (довгоносики,
жужелиці, ковалики, стафілініди).
Ми проаналізували наявність видів різних
зоогеографічних елементів грунтової мезофауни в соснових лісах регіону
дослідження, що вісвітлено у таблиці 1.
Встановлено 7 зоогеографічних елементів:
європейський (35,1%), транспалеарктичний (30,8%), європейсько-сибірський
(21,3%), голарктичний, космополітний і европейсько-середземноморський (у цілому
12,8%). Домінуючими є види європейські, транспалеарктичні та
європейсько-сибірські, що відображено у рисунку 2. Встановили, що серед них в
сосняках переважають види з більш вузькими ареалами.
В березових лісах переважають транспалеарктичні
(38,6) і європейські (53,8%) види, що продемонстровано у рисунку 3.
При оцінці чисельності грунтової мезофауни, яка
належать по певної трофічної групи, виявилося, що кількість фітофагів в
березняках чорничних становить 18,6% значно нижча, ніж в сосняках мохових,
сапрофагів - 23,4%, а зоофагів - 53,5, найнижчі величини мають некрофаги -
4,5%, що продемонстровано у рисунку 4.
У сосняках по чисельності і зоомасі переважають
зоофаги (72,5%), набагато менше фітофагів (13,7%) і детритофагів (9,9%),
незначна чисельність копрофагів (3,8%), що відображено в рисунку 5. Бідність
сосняків сапрофагами можна пояснити відсутністю в підстилці листяного опаду,
незначним змістом у ґрунті гумусу і незначною придатністю соснової хвої для
живлення ґрунтових тварин.
На основі проведених досліджень зроблені наступні
висновки:
. Встановлений фауністичний склад представників
ґрунтової мезофауни у домінуючих лісах Черемського природного заповідника.
Всього виявлено 106 видів ґрунтової мезофауни, які належать до наступних
таксономічних груп: Олігохети (Oligochaeta), Комахи (Insecta), Багатоніжки
(Myriapoda), Ракоподібні (Crustacea), Павукоподібні (Araneamorpha), Молюски
(Mollusca).
. Проаналізовано екологічні особливості поширення
грунтової мезофауни у різних типах лісу Черемського природного заповідника,
зокрема, у сосняках лишайникових (Pineum cladinosum), сосняках сфагнових
(Pineum sphagnosum), сосняках чорничних (Pineum myrtillosum), сосняках мохових
(Pineum pleuroziosum), а також березових лісах.
. Встановлено кількісний склад і біомасу
представників ґрунтової мезофауни у соснових та березових лісах Черемського
природного заповідника.
. Фауністичний склад грунтової мезофауни у
соснових лісах Черемського природного заповідника включає представників із 5
класів, 8 рядів, 14 родин, 42 види.
Домінуючими представниками ґрунтової мезофауни
соснових лісів є комахи, серед яких практично у всіх типах сосняків переважають
твердокрилі, жужелиці та стафіленіди.
Малочисельними є олігохети - 4,4 екз/м2,
які слугують індикаторами соснових типів лісів, де їх чисельність варіює від
0,8 до 5,6 % від чисельності всіх безхребетних.
Досліджено, що найменше представленою є ґрунтова
мезофауна у сосняках лишайникових (Pineum cladinosum).
. Найкраще розвинутою є ґрунтова мезофауна у
березових лісах. Фауністичний склад грунтової мезофауни у березових лісах
Черемського природного заповідника включає представників із 5 класів, 11 рядів,
17 родин, 64 видів.
Середня чисельність грунтової мезофауни в
березняках, досить висока і складає 244,0-253,7 екз/м2.
Дощові черви та комахи займають по чисельності
домінуюче становище в березових лісах. Серед комах преважають твердокрилі і
двокрилі. Перші представлені 8 родинами; 4 із них домінують (довгоносики,
жужелиці, ковалики, стафілініди).
4.Проаналізували наявність видів різних
зоогеографічних елементів грунтової мезофауни в соснових лісах регіону
дослідження. Встановлено 7 зоогеографічних елементів: європейський (35,1%),
транспалеарктичний (30,8%), європейсько-сибірський (21,3%), голарктичний,
космополітний і европейсько-середземноморський (у цілому 12,8%). Домінуючими є
види європейські, транспалеарктичні та європейсько-сибірські. Встановили, що
серед них в сосняках переважають види з більш вузькими ареалами. В березових
лісах переважають транспалеарктичні (38,6) і європейські (53,8%) види.
7. При оцінці чисельності грунтової мезофауни,
яка належать по певної трофічної групи, виявилося, що кількість фітофагів в
березняках чорничних становить 18,6% значно нижча, ніж в сосняках мохових,
сапрофагів - 23,4%, а зоофагів - 53,5, найнижчі величини мають некрофаги -
4,5%. У сосняках по чисельності і зоомасі переважають зоофаги (72,5%), набагато
менше фітофагів (13,7%) і детритофагів (9,9%), незначна чисельність копрофагів
(3,8%). Бідність сосняків сапрофагами можна пояснити відсутністю в підстилці
листяного опаду, незначним змістом у ґрунті гумусу і незначною придатністю
соснової хвої для живлення ґрунтових тварин.
ДОПОВІДЬ 2
Шановні присутні. На Ваш
розгляд пропонується
дипломна робота на тему:„ ЕКОЛОГО-ФАУНІСТИЧНА
ХАРАКТЕРИСТИКА МЕЗОФАУНИ Черемського
природного заповідника” (РИС. 1)
Мезофауна є важливим компонентом біогеоценозів.
Вона займає ведуче місце в зоомасі в складі ценозу, істотно впливає на процеси
трансформації органічних решток, міграції хімічних елементів в горизонтах
ґрунтового профілю, ґрунтоутворення, стимулювання біологічних процесів та
сукцесії організмів в біогеоценозах ґрунту. Без обліку мезофауни населення
ґрунту неможливо скласти загальний кадастр тваринного світу. Разом з тим,
мезофауна ґрунту включає значну кількість лісових і сільськогосподарських
шкідників, а тому має велике практичне значення.
Серед представників мезофауни ґрунтових
безхребетних тварин є багато стенобіонтних видів. Власне вони відповідають
вимогам індикації стану оточуючого середовища і можуть служити об‘єктами
моніторингу. Розв‘язати завдання в напрямку біоіндикаційних цілей можна лише на
основі дослідження угрупування ґрунтових безхребетних в цілому, в тому числі,
представників мезофауни.
Актуальність дослідження зумовлена недостатнім
рівнем відомостей про фауністичний склад, особливості екології та поширення
ґрунтової мезофауни Черемського природного заповідника.
Мета роботи: вивчити еколого-фауністичні
особливості ґрунтової мезофауни району дослідження (РИС. 2)
В роботі ставились такі завдання:
. Проаналізувати літературні джерела та
ознайомитись з історією вивчення ґрунтової мезофауни.
. Розглянути фізико-географічні умови Черемського
природного заповідника.
. Встановити фауністичний склад ґрунтової
мезофауни у районі дослідження.
. Вивчити екологію ґрунтової мезофауни у
Черемському природному заповіднику.
. Виявити особливості поширення мезофауни у
районі вивчення.
Об’єктом дослідження є ґрунтова мезофауна
Черемського природного заповідника (РИС. 3.)
Предметом дослідження є фауна, екологія,
особливості поширення ґрунтової мезофауни у районі дослідження.
Наукова новизна дослідження. Вперше вивчено
видовий склад, особливості екології та хорології грунтової мезофауни у
Черемському природному заповіднику.
Практичне значення дипломної роботи полягає в
тому, що його результати можуть бути використанні у професійній діяльності
педобіологів, ґрунтознавців, агрономів, працівників сільського господарства та
лісництва, а також у навчальному процесі. Дані роботи можна використовувати
також при проведенні інтродукції та акліматизації окремих видів у відповідності
до типів ґрунтів.
Матеріал для дипломної
роботи був зібраний протягом 2008-2009 рр. у Черемському природному заповіднику
(РИС. 4) Було відібрано 110 проб. Для вивчення видового складу грунтової мезофауни ми
користувалися методиками М.С. Гілярова та К.К. Фасулатті.
Ми проаналізували наявність видів різних
зоогеографічних елементів грунтової мезофауни в соснових лісах району
дослідження, що вісвітлено у таблиці . (РИС. 5.)
Встановлено 7 зоогеографічних елементів:
європейський (35,1%), транспалеарктичний (30,8%), європейсько-сибірський
(21,3%), голарктичний, космополітний і европейсько-середземноморський (у цілому
12,8%). Домінуючими є види європейські, транспалеарктичні та
європейсько-сибірські (РИС. 6). Встановили, що серед них в сосняках переважають
види з більш вузькими ареалами.
В березових лісах переважають транспалеарктичні
(38,6) і європейські (53,8%) види (РИС. 7)
При оцінці чисельності грунтової мезофауни, яка
належать по певної трофічної групи, виявилося, що кількість фітофагів в
березняках чорничних становить 18,6% значно нижча, ніж в сосняках мохових,
сапрофагів - 23,4%, а зоофагів - 53,5, найнижчі величини мають некрофаги - 4,5%
(РИС. 8)
У сосняках по чисельності і зоомасі переважають
зоофаги (72,5%), набагато менше фітофагів (13,7%) і детритофагів (9,9%),
незначна чисельність копрофагів (3,8%) (РИС. 9). Бідність сосняків сапрофагами
можна пояснити відсутністю в підстилці листяного опаду, незначним змістом у
ґрунті гумусу і незначною придатністю соснової хвої для живлення ґрунтових
тварин.
На основі проведених
досліджень зроблені наступні висновки: (РИС. 10)
1. Встановлений фауністичний склад представників
ґрунтової мезофауни у домінуючих лісах Черемського природного заповідника.
Всього виявлено 110 видів ґрунтової мезофауни, які належать до наступних
таксономічних груп: Комахи (Insecta), Багатоніжки (Myriapoda), Олігохети
(Oligochaeta), Молюски (Mollusca), Ракоподібні (Crustacea), Павукоподібні
(Araneamorpha).
. Проаналізовано екологічні особливості поширення
грунтової мезофауни у різних типах лісу Черемського природного заповідника,
зокрема, у сосняках лишайникових (Pineum cladinosum), сосняках сфагнових
(Pineum sphagnosum), сосняках чорничних (Pineum myrtillosum), сосняках мохових
(Pineum pleuroziosum), а також березових лісах.
. Встановлено кількісний склад і біомасу
представників ґрунтової мезофауни у соснових та березових лісах Черемського
природного заповідника.
. Фауністичний склад грунтової мезофауни у
соснових лісах Черемського природного заповідника включає представників із 5
класів, 8 рядів, 14 родин, 42 види.
Домінуючими представниками ґрунтової мезофауни
соснових лісів є комахи, серед яких практично у всіх типах сосняків переважають
твердокрилі, жужелиці та стафіленіди. Малочисельними є олігохети - 4,4 екз/м2,
які слугують індикаторами соснових типів лісів, де їх чисельність варіює від
0,8 до 5,6 % від чисельності всіх безхребетних.
Досліджено, що найменше представленою є ґрунтова
мезофауна у сосняках лишайникових (Pineum cladinosum).
. Найкраще розвинутою є ґрунтова мезофауна у
березових лісах. Фауністичний склад грунтової мезофауни у березових лісах
Черемського природного заповідника включає представників із 5 класів, 11 рядів,
17 родин, 64 видів. Середня чисельність грунтової мезофауни в березняках,
досить висока і складає 244,0-253,7 екз/м2.
Дощові черви та комахи займають по чисельності
домінуюче становище в березових лісах. Серед комах преважають твердокрилі і
двокрилі. Перші представлені 8 родинами; 4 із них домінують (довгоносики,
жужелиці, ковалики, стафілініди).
5.При оцінці чисельності грунтової мезофауни, яка
належать по певної трофічної групи, виявилося, що кількість фітофагів в
березових лісах становить 18,6%, сапрофагів - 23,4%, а зоофагів - 53,5,
найнижчі величини мають некрофаги - 4,5%. У соснових лісах по чисельності і зоомасі
переважають зоофаги (72,5%), набагато менше фітофагів (13,7%) і детритофагів
(9,9%), незначна чисельність копрофагів (3,8%). Бідність сосняків сапрофагами
можна пояснити відсутністю в підстилці листяного опаду, незначним змістом у
ґрунті гумусу і незначною придатністю соснової хвої для живлення ґрунтових
тварин.
6.Проаналізували наявність видів різних
зоогеографічних елементів грунтової мезофауни в соснових лісах регіону
дослідження. Встановлено 7 зоогеографічних елементів: європейський (35,1%),
транспалеарктичний (30,8%), європейсько-сибірський (21,3%), голарктичний,
космополітний і европейсько-середземноморський (у цілому 12,8%). Домінуючими є
види європейські, транспалеарктичні та європейсько-сибірські. Встановили, що
серед них в сосняках переважають види з більш вузькими ареалами. В березових
лісах переважають транспалеарктичні (38,6) і європейські (53,8%) види.
ДОПОВІДЬ 3
Шановні присутні на Ваш розгляд
пропонується дипломна робота на тему : «Еколого-фауністичний аналіз грунтової
мезофауни Рожищенського району»
Ґрунт - природне біокосне тіло, поряд з такими
його складовими, як тверда, рідка і газоподібна фази, містить і живу частину.
До складу ґрунтової біоти входять представники всіх царств живої природи.
Ґрунтове населення не тільки живе в ґрунті,
використовуючи його як середовище існування, але й активно трансформує його.
Тому цілком закономірне твердження, що ґрунт - це середовище існування й
одночасно продукт діяльності живих істот, що його населяють.
Ґрунтова біота - найбільш динамічна ланка екосистеми,
яка зазнає періодичних змін і, у той же час, що володіє певним гомеостазом
завдяки наявності механізмів регуляції і контролю.
Грунтові біоценози представлені
найрізноманітнішими безхребетними тваринами, а саме: найпростішими
(саркодовими, джгутиковими, інфузоріями), нематодами, коловертками, олігохетами
(люмбрицідами і енхитреїдами), черевоногими молюсками, хеліцеровими (павуками,
кліщами, косариками, псевдоскорпіонами), ракоподібними, багатоніжками,
комахами. При урахуванні особливостей життя і впливу на грунт тварин різних
розмірів їх поділяють на три групи: мікрофауну, мезофауну та мегафауну.
Мезофауна є важливим компонентом біогеоценозів.
Вона займає ведуче місце в зоомасі в складі ценозу, істотно впливає на процеси
трансформації органічних решток, міграції хімічних елементів в горизонтах
ґрунтового профілю, ґрунтоутворення, стимулювання біологічних процесів та
сукцесії організмів в біогеоценозах ґрунту. Без обліку мезофауни населення
ґрунту неможливо скласти загальний кадастр тваринного світу. Разом з тим,
мезофауна ґрунту включає значну кількість лісових і сільськогосподарських
шкідників, а тому має велике практичне значення.
Серед представників мезофауни ґрунтових
безхребетних тварин є багато стенобіонтних видів. Власне вони відповідають вимогам
індикації стану оточуючого середовища і можуть служити об‘єктами моніторингу.
Розв‘язати завдання в напрямку біоіндикаційних
цілей можна лише на основі дослідження угрупування ґрунтових безхребетних в
цілому, в тому числі, представників мезофауни.
Актуальність дослідження зумовлена недостатнім
рівнем відомостей про фауністичний склад, особливості екології та поширення
ґрунтової мезофауни Рожищенського району.
Мета роботи: вивчити еколого-фауністичні
особливості ґрунтової мезофауни району дослідження.
В роботі ставились такі завдання:
. Проаналізувати літературні джерела та
ознайомитись з історією вивчення ґрунтової мезофауни.
. Розглянути фізико-географічні умови
Рожищенського району
. Встановити фауністичний склад ґрунтової
мезофауни у районі дослідження.
. Вивчити екологію ґрунтової мезофауни у
Рожищенському районі.
. Виявити особливості поширення мезофауни у
районі вивчення.
Об’єктом дослідження є ґрунтова мезофауна
Рожищенського району. Предметом дослідження є фауна, екологія, особливості
поширення ґрунтової мезофауни у районі дослідження.
Наукова новизна дослідження. Вперше вивчено
видовий склад, особливості екології та хорології грунтової мезофауни у
Рожищенському районі.
Практичне значення дипломної роботи полягає в
тому, що його результати можуть бути використанні у професійній діяльності
педобіологів, ґрунтознавців, агрономів, працівників сільського господарства та
лісництва, а також у навчальному процесі. Дані роботи можна використовувати
також при проведенні інтродукції та акліматизації окремих видів у відповідності
до типів ґрунтів.
Матеріал для дипломної
роботи був зібраний протягом 2009-2010 рр. у Рожищенському районі. Було
відібрано 120 проб. Місця відбору проб вказані на карті-схемі.
Для вивчення видового складу грунтової мезофауни
ми користувалися методиками М. С. Гілярова та К. К. Фасулатті.
На основі проведених досліджень зроблені настпні
висновки:
. Встановлений фауністичний склад представників
ґрунтової мезофауни у домінуючих лісах Рожищенського району. Всього виявлено
110 видів ґрунтової мезофауни, які належать до наступних таксономічних груп:
Комахи (Insecta), Багатоніжки (Myriapoda), Олігохети (Oligochaeta), Молюски
(Mollusca), Ракоподібні (Crustacea), Павукоподібні (Araneamorpha).
. Проаналізовано екологічні особливості поширення
грунтової мезофауни у різних типах лісу Рожищенського району, зокрема, у
сосняках лишайникових (Pineum cladinosum), сосняках сфагнових (Pineum
sphagnosum), сосняках чорничних (Pineum myrtillosum), сосняках мохових (Pineum
pleuroziosum), а також березових лісах.
. Встановлено кількісний склад і біомасу
представників ґрунтової мезофауни у соснових та березових лісах Рожищенського
району.
. Фауністичний склад грунтової мезофауни у
соснових лісах Рожищенського району включає представників із 5 класів, 8 рядів,
14 родин, 42 види.
Домінуючими представниками ґрунтової мезофауни
соснових лісів є комахи, серед яких практично у всіх типах сосняків переважають
твердокрилі, жужелиці та стафіленіди. Малочисельними є олігохети - 4,4 екз/м2,
які слугують індикаторами соснових типів лісів, де їх чисельність варіює від
0,8 до 5,6 % від чисельності всіх безхребетних.
Досліджено, що найменше представленою є ґрунтова
мезофауна у сосняках лишайникових (Pineum cladinosum).
. Найкраще розвинутою є ґрунтова мезофауна у
березових лісах. Фауністичний склад грунтової мезофауни у березових лісах
Рожищенського району включає представників із 5 класів, 11 рядів, 17 родин, 64
видів.Середня чисельність грунтової мезофауни в березняках, досить висока і
складає 244,0-253,7 екз/м2.
Дощові черви та комахи займають по чисельності
домінуюче становище в березових лісах. Серед комах преважають твердокрилі і
двокрилі. Перші представлені 8 родинами; 4 із них домінують (довгоносики,
жужелиці, ковалики, стафілініди).
7.При оцінці чисельності грунтової мезофауни, яка
належать по певної трофічної групи, виявилося, що кількість фітофагів в
березових лісах становить 18,6%, сапрофагів - 23,4%, а зоофагів - 53,5,
найнижчі величини мають некрофаги - 4,5%. У соснових лісах по чисельності і
зоомасі переважають зоофаги (72,5%), набагато менше фітофагів (13,7%) і
детритофагів (9,9%), незначна чисельність копрофагів (3,8%). Бідність сосняків
сапрофагами можна пояснити відсутністю в підстилці листяного опаду, незначним
змістом у ґрунті гумусу і незначною придатністю соснової хвої для живлення
ґрунтових тварин.
8.Проаналізували наявність видів різних
зоогеографічних елементів грунтової мезофауни в соснових лісах регіону
дослідження. Встановлено 7 зоогеографічних елементів: європейський (35,1%),
транспалеарктичний (30,8%), європейсько-сибірський (21,3%), голарктичний,
космополітний і европейсько-середземноморський (у цілому 12,8%). Домінуючими є
види європейські, транспалеарктичні та європейсько-сибірські. Встановили, що
серед них в сосняках переважають види з більш вузькими ареалами. В березових
лісах переважають транспалеарктичні (38,6) і європейські (53,8%) види.