Лютеинизирующий и фолликулостимулирующий гормоны. Физиологическая роль и механизмы регуляции
Реферат
по
эндокринологии.
Лютеинизирующий
и фолликулостимулирующий гормоны. Физиологическая роль и механизмы регуляции.
Выполнила студентка Локтионова А.
Преподаватель Смирнова О.В.
Оглавление
Введение
Гонадотропинов
рилизинг-гормон
Эффекты гонадотропинов в
пределах женской репродуктивной системы
Механизм действия
гонадотропинов
Гипофизарный контроль
образования гормонов яичников
Нарушение регуляции
гонадотрофов ассоциировано с рядом заболеваний
Методы изучения функций
гонадотрофов
Дифференциальная активация
сигнальных каскадов пульсирующим или постоянным выбросом ГнРГ
Декодирование импульсов ГнРГ
посредством ERK-сигнального каскада
Эпигенетическая регуляция
функций гонадотрофов
Заключение
Список литературы
Введение
В последнее время были выявлены новые особенности работы гонадотрофов
передней доли гипофиза, касающиеся различных механизмов продукции и секреции
лютеинизирующего (ЛГ) и фолликулостимулирующего (ФСГ) гормонов. Эти открытия
могут найти применение в терапевтических методах лечения заболеваний, связанных
с женским репродуктивным здоровьем.
У зрелых гонадотрофов выявлена связь дифференциальной экспрессии гена,
кодирующего β-субъединицы гонадотропинов, с активацией и/или стабилизацией
важных регуляторных белков и транскрипционных факторов. В ходе последних
исследований возникла идея того, что внутриклеточные процессы, влияющие на
модификации гистонов, играют важную роль как в развитии гонадотрофов, так и в
экспрессии генов, кодирующих субъединицы гонадотропинов. Динамика секреции
гонадотрофов контролируется гипоталамическим нейропептидом -
гонадотропин-рилизинг гормоном (ГнРГ) потенциально через способность ГнРГ
активировать несколько различных сигнальных каскадов к гонадотрофам.
Таким образом, глубокое проникновение в клеточные процессы является
ключом к физиологии гонадотрофов и к пониманию причин неправильной секреции
гонадотропинов, приводящей к таким заболеваниям как гипоталамическая аменорея и
синдром поликистозных яичников (СПКЯ).
Пульсирующий характер синтеза и секреции круциален для многих
физиологических процессов, потому как нарушение эпизодических выбросов гормона
часто приводит к клиническим расстройствам. Колебания секреции ЛГ и ФСГ
находятся под контролем «пульсирующего» ГнРГ. Варианты в паттерне импульсации
ГнРГ ассоциированы с дивергирующей секрецией ЛГ и ФСГ, обеспечивая механизм
регуляции действия двух различных гормонов, выделяемых схожими типами клеток
(гонадотрофами), одним гипоталамическим нейропептидом. Вероятно, возможность
такой регуляции опирается на способность гонадотрофов «расшифровывать» разные
паттерны секреции ГнРГ. Исследования достаточно хорошо изученных гонадотрофов
типа αТ3-1 и LβТ2 показали, что как влияние
активации и/или стабилизации регуляторных белков, так и эпигенетических
процессов одинаково важно для поддержания динамики ответа гонадотрофов на
воздействие ГнРГ.
Гонадотропинов рилизинг-гормон
Стимулирует
высвобождение и ЛГ, и ФСГ. Относится к семейству декапептидов, высоко
консервативных на протяжении эволюции позвоночных. К настоящему времени у
видов, отличных от млекопитающих, было описано 15 различных вариантов.
Примечательно, что пептидная последовательность ГнРГ сопровождается
последовательностью ассоциированного полипептида варьирующей длины. До недавнего
времени полагали, что ГнРГ млекопитающих кодируется одним геном. В настоящее
время имеются доказательства существования второго гена. Первый ген ГнРГ (ГнРГ-I)
кодирует белок-предшественник из 92 аминокислот, который затем после
протеолитического расщепления даёт декапептид ГнРГ и 56-аминокислотный
ассоциированный пептид. Второй ген ГнРГ (ГнРГ-II) кодирует высококонсервативный
декапептид, отличающийся от ГнРГ-I по 3 аминокислотным остаткам. Геномная
структура этого гена сходна с таковой ГнРГ-I, но локализован на другой
хромосоме (хромосоме 20 вместо 8). Ген ГнРГ-II кодирует ассоциированный пептид,
который на 50% длиннее ассоциируемого пептида, кодируемого ГнРГ-I (84 и 56
аминокислотных остатков соответственно). Функция ГнРГ-II в настоящее время неизвестна.
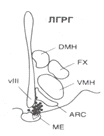
У
человека значительная часть ГнРГ-нейронов локализуется в медиобазальном
гипоталамусе и аркуатном ядре (см. Рис.1) ГнРГ-нервные окончания,
проецирующиеся на портальную систему сосудов, расположены преимущественно на
латеральной части срединного возвышения, хотя наблюдается также и медиальное
распределение. Нейроны, содержащие ГнРГ-II, также обнаруживаются в
гипоталамусе, но в областях, отличных от тех, которые продуцируют ГнРГ- I.
На
рисунке: DMH - дорзомедиальное гипоталамическое ядро; FX -
свод; VMH - вентромедиальное гипоталамическое ядро; ARC -
аркуатное ядро; ME - срединное возвышение; vIII- третий
желудочек.
Эффекты гонадотропинов в пределах женской репродуктивной
системы. Механизм действия гонадотропинов
Гормоны ЛГ и ФСГ действуют на яичник путём исходного связывания с
высокоаффинными специфическими мембранными рецепторами с низкой ёмкостью. ФСГ
инициирует свои эффекты благодаря связыванию с сопряжённым с G-белком рецептором, содержащим семь
трансмембранных доменов. Рецептор представляет собой гликопротеин с
предсказанной молекулярной массой около 75000, кодируемый одним геном,
картируемым на 2 хромосоме генома человека. Хотя рецептор ЛГ картирован на той
же хромосоме, он не локализован в той же области хромосомы. Белок рецептора ФСГ
человека кодируется открытой рамкой считывания из 2085 нуклеотидов, которая
даёт зрелый белок из 678 аминокислот с предсказанной молекулярной массой около
75000. Экспрессия рецептора высокоспецифична, поскольку его удаётся обнаружить
только в клетках Сертоли семенника и гранулёзных клеток яичника. Связывание ФСГ
приводит к образованию цАМФ и мобилизации кальция. Не ясно, является ли
мобилизация кальция следствием повышения образования цАМФ, или независимым от
цАМФ феноменом. Известно, однако, что цАМФ активирует протеинкиназу А, которая
в свою очередь индуцирует фосфорилирование нескольких нижележащих белков,
участвующих в регуляции экспрессии цАМФ-чувствительных генов.
Рецептор ЛГ также сопряжён с G-белком рецептором, содержащим семь трансмембранных доменов. Его
аминокислотная последовательность проявляет значительную гомологию с рецептором
ФСГ, особенно в трансмембранных доменах. Зрелый белок представляет собой
полипептид, содержащий 674 аминокислотных остатка. Его короткий внутриклеточный
домен содержит ряд потенциальных сайтов фосфорилирования. Интересно, что, хотя
активация рецептора ЛГ вызывает явное увеличение уровня цАМФ, внутриклеточный
домен рецептора содержит слабый консенсус для фосфорилирования, опосредуемого
зависимой от цАМФ протеинкиназой.
Гипофизарный контроль образования гормонов яичников
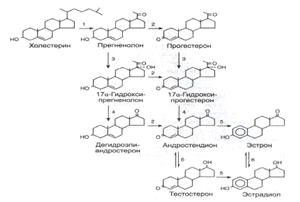
Рис.2. Биосинтез стероидов в яичнике. 1: комплекс фермента, отщепляющего
боковую цепь холестерина; 2: 3β-гидроксистероиддегидрогеназа; 3: 17α-гидроксилаза; 4: 17,20-лиаза; 5:
ароматаза; 6: 17β-стероиддегидрогеназа.[2]
Под контролем ЛГ и ФСГ находится продукция стероидов яичников. Также,
эксперименты на грызунах показали, что в зависимости от уровня гормона
пролактина последний может либо ингибировать, либо поддерживать стимулирующее
действие ЛГ на стероидогенез в яичниках посредством модуляции количества
рецепторов ЛГ. А вот гормон роста, напротив, у крыс облегчает действие ФСГ
посредством механизма, включающего местную продукцию ИФР-1.
Секреция прогестерона может стимулироваться и ЛГ, и ФСГ, секреция
андрогенов увеличивается только под воздействием ЛГ. Образование эстрадиола
зависит от обоих гонадотропинов. В то время как ЛГ стимулирует продукцию
андрогенов, ФСГ прямо активирует ферментный комплекс ароматазы, который
катализирует превращение андрогенов в эстрогены.
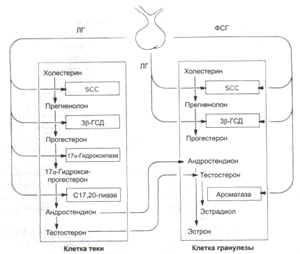
Рис.3. Контроль продукции эстрогенов, прогестерона и
андрогенов в яичниках посредством ЛГ и ФСГ. ЛГ действует на клетки теки и
гранулёзы; ФСГ - только на клетки гранулёзы. [3]
Рецепторы ЛГ обнаружены в стероидогенных клетках стромы, жёлтого тела и
клетках теки фолликулов; все они продуцируют прогестерон и андрогены. Когда
фолликулы становятся антральными, клетки гранулёзы приобретают рецепторы к ЛГ.
Поскольку эти клетки содержат ограниченную 17,20-лиазную активность, их
экспозиция с ЛГ приводит преимущественно к повышению продукции прогестерона и
небольшой, если она вообще есть, продукции андрогенов. Таким образом, хотя
клетки гранулёзы наделены существенной ароматазной активностью, ЛГ не может
стимулировать образование эстрогенов, если только андрогены не обеспечиваются
клетками другого типа. То же самое относится и к ФСГ. Его рецепторы
локализованы исключительно на клетках гранулёзы, которые отвечают на ФСГ
образованием эстрогенов, если они адекватно снабжены андрогенами.
Эти взаимодействия послужили основой для выдвижения гипотезы две
клетки - два гонадотропина для объяснения гонадотропного контроля
стероидогенеза яичников (См. рис.3).
Согласно этой гипотезе, тека фолликула под влиянием ЛГ продуцирует
андрогены, которые после диффузии к компартменту гранулёзных клеток фолликула
превращаются в эстрогены посредством поддерживаемой ФСГ реакции ароматизации.
Нарушение регуляции гонадотрофов ассоциировано с рядом
заболеваний
Тесная взаимосвязь между секрецией ГнРГ и продукцией гонадотропинов
доказана на примере больных с синдромом Кальмана, у которых недостаток ГнРГ
выражается низким уровнем синтеза гонадотропинов, отсутствием полового
созревания, бесплодием. Таким образом, контроль биосинтеза и секреции ЛГ и ФСГ
гонадотропин-рилизинг гормоном зависит от паттерна доставки ГнРГ к гонадотрофам
передней доли гипофиза.
Напротив, снижение частоты выброса ГнРГ, характерное для лютеальной и
ранней фолликулярной стадий менструального цикла, проявляется преимущественной
секрецией ФСГ, играющему важную роль в процессах выбора доминантного фолликула.
Таким образом, ответ гонадотрофов на ГнРГ (путём дивергенции синтеза и
секреции ЛГ и ФСГ) крайне чувствителен к паттерну его выброса. Это иллюстрирует
синдром поликистозных яичников, самая распространённая причина бесплодия у
женщин репродуктивного возраста. Патогенез этой болезни до конца неясен, но
отличительным признаком данного заболевания является нарушение менструального
цикла, связанное с повышением содержания ЛГ в сыворотке крови и снижением
содержания ФСГ, и это приводит к повышению продукции андрогенов клетками теки
яичников. Это изменение динамики гонадотропинов приводит к повышенной
нейрональной активности гипоталамуса, которая проявляется увеличением частоты
выброса ГнРГ. Напротив, гипоталамическая аменорея у женщин связана с низкой
частотой выброса ГнРГ.
Методы изучения функций гонадотрофов
Передний гипофиз - совокупность различных типов клеток, каждый из которых
вырабатывает определённый гормон. Несмотря на то, что гонадотрофы составляют
лишь 5-10% от числа клеток гипофиза, они играют ведущую роль в становлении и
поддержании фертильности. Учитывая малое количество этих клеток в гетерогенном
гипофизе, изучение гонадотрофов в первичных культурах клеток гипофиза является
трудной задачей. Поэтому культивирование клеточных линий αТ3-1 и LβТ2 - типов даёт базу для изучения
молекулярных и клеточных процессов, лежащих в основе синтеза и секреции ЛГ и
ФСГ.
Важно отметить, что искусственно поддерживаемые клеточные линии по
характеристикам отличны от зрелых гонадотрофов функционирующего гипофиза, и
есть возможность, что они будут отражать лишь немногую часть всех свойств,
характерных для данного клеточного типа. Чтобы преодолеть эти ограничения,
исследователи разработали методику выделения, идентификации и очистки
гонадотрофов из гипофиза трансгенных животных.
Дифференциальная активация сигнальных каскадов пульсирующим
или постоянным выбросом ГнРГ
Гонадотропин-рилизинг гормон - ГнРГ - воздействует на гонадотрофы
гипофиза посредством рецепторов, сопряжённых с G-белками. Последствия этой сигнализации зависят от частоты
пульсации ГнРГ; но механизмы декодирования данной частоты остаются не до конца
выясненными. Важную роль в них играют внеклеточные сигнал-регулирующие киназы (ERK), митоген-активирующие протеинкиназы
(МАРК), и другие посредники.
Как было сказано выше, ГнРГ выделяется гипоталамусом в виде коротких
импульсов и посредством семи трансмембранных рецепторов воздействует на
гонадотрофы, стимулируя фосфолипазу С, мобилизуя Са2+ и активируя
изозимы протеинкиназы С. Это ведёт к активации МАРК и запуску МАР-киназного
каскада, а также стимулирует Са2+-эффекторы (такие как кальмодулин).
Последние, в свою очередь, опосредуют эффекты ГнРГ на гонадотрофы, влияя на
экспрессию множества генов, включая гены субъединиц ЛГ и ФСГ, и на секрецию
гонадотропинов.
Эффекты ГнРГ частотно-зависимы. Постоянная секреция гипоталамусом ГнРГ
подавляет секрецию и ЛГ, и ФСГ, в то время как возобновление пульсирующей
секреции этого гормона индуцирует нормальное физиологическое выделение гонадотропинов
in vivo. Такая десенситизация ГнРГ-индуцируемой секреции
гонадотропинов используется в терапевтических целях: пульсирующее введение
агонистов ГнРГ может повысить количество циркулирующих в крови гонадотропинов и
половых стероидов, что в свою очередь повышает возможность зачатия (такая
методика используется, например, при гиперстимуляции яичников, предшествующей
искусственному оплодотворению). Напротив, устойчивое присутствие агонистов
ГнРГ, вводимых в организм, снижает секрецию стероидов, что лежит в основе
лечения форм рака, зависимых от стероидных гормонов.
Когда частота пульсации ГнРГ возрастает до физиологического уровня,
увеличивается влияние этого гормона на экспрессию генов, кодирующих β-субъединицы ЛГ и ФСГ а также тех
генов, которые кодируют рецепторы к ГнРГ (ГнРГР). Но если частота станет
превышать физиологические значения, транскрипция снизится. Вычислительные
модели показывают, что такие колоколообразные амплитудно-частотные
взаимоотношения требуют механизмов обратной связи, которые могут включать
отрицательную регуляцию ГнРГ, индукцию RGS-2 (регулятор сигнализации G-белка),
ингибирование Са2+-каналов кальмодулин-зависимым G-белком Kir/Gem,
или индукцию фосфатазы MAP-киназы.
Но быстрой десенситизации ГнРГ-рецепторов типа 1 у человека не происходит, что
говорит о присутствии некоего механизма, предупреждающего такое развитие
событий. Этот механизм заключается в отсутствии у этих рецепторов С-концевого
участка, который опосредует фосфорилирование, связывание аррестина и
десенситизацию многих других трансмембранных рецепторов схожего типа.
Альтернативный механизм расшифровки пульсирующих сигналов ГнРГ включает
взаимодействия между транскрипционным фактором Egr-1 (белок раннего ответа) и
корегулятором (Nab-2) в промоторе β-субьединицы ЛГ. Согласно этой модели,
низкая частота пульсации ГнРГ вызывает непродолжительную экспрессию Egr-1
(вызванную экспрессией Nab-2), что ингибирует экспрессию гена β-субьединицы ЛГ. В то же время,
повышение частоты пульсации ГнРГ вызовет более устойчивое усиление экспрессии
Egr-1, погасит Nab-2 и индуцирует экспрессию гена β-субьединицы ЛГ (хотя и неясно,
происходит ли это in vivo). Сходные взаимодействия происходят
в рамках промотора ФСГ (между регулятором транскрипции c-Fos и корегулятором
TGIF), являясь основой экспрессии гена β-субьединицы соответствующего гормона
при низкой частоте пульсации ГнРГ. Некоторые исследователи предлагают ещё один
вариант декодирования, считая что роль могут играть CREB (фактор, связывающий
цAMФ-респонсивный элемент) и цАМФ-индуцибельный репрессор. В этом случае
высокочастотные импульсы ГнРГ активируют цАМФ-индуцибельный репрессор, вызывая
подавление транскрипции путём конкурирования за сайт CRE (цAMФ-респонсивный
элемент) в промоторе гена β-субъединицы ФСГ.
Итак, вышеприведенные данные описывают 2 возможных механизма
декодирования импульсов ГнРГ: частота пульсации либо отражается механизмами
обратной связи и запускает сигнальные каскады в цитоплазме, либо декодируется
уровнем транскрипции и экспрессии определённых генов. Исследовать первый из
предполагаемых путей, используя обычные методы, оказалось довольно сложно.
Исследователи изучают возможность декодирования импульсов ГнРГ посредством ERK-сигнального каскада.
Декодирование импульсов ГнРГ посредством ERK-сигнального каскада
Как и многие другие семи-доменные трансмембранные рецепторы, ГнРГР
запускает Raf/MEK/ERK-комплекс.- консервативный сигнальный белок, активируемый
Raf-серин/треонин киназами. Raf активирует MAPK/ERK
киназу (MEK)1/2 - протеинкиназу двойной
специфичности - которая затем активирует ERK1/2 фосфорилированием. Raf
активируется изозимами протеинкиназы С (их активность как раз индуцирует ГнРГ,
связываясь с ГнРГР).
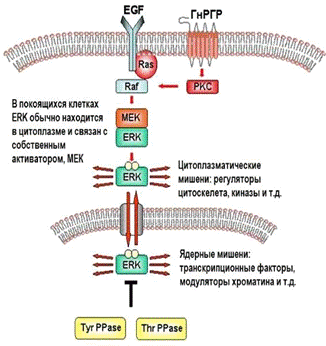
Рис.4 Общая схема функционирования Raf/MEK/ERK-комплекса[4]
Важнейшую роль в регуляции ERK
играет т.н. TEY-участок (Thr-Glu-Tyr). Его
фосфорилирование активирует ERK и
освобождает от цитоплазматических «якорей», позволяя войти в ядро клетки. В
ядре ERK контролирует экспрессию генов,
фосфорилируя факторы транскрипции. Таким образом, ГнРГ-активированные ERK1/2 являются медиаторами
декодирования пульсации ГнРГ: они трансформируют частоту пульсации в
регулирование транскрипции генов β-субъединиц ЛГ и ФСГ. (Рис.2)
ГнРГ также активирует МКР из семейства протеинкиназ двойной специфичности
(DUSP), и вычислительные модели иллюстрируют возможную причастность
МАР-киназных каскадов и активируемых фосфатаз к декодированию импульсов ГнРГ.
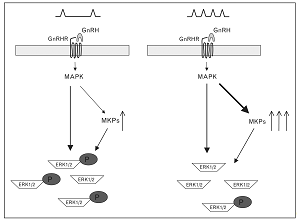
Рис.5 Зависимость количества активированных молекул ERK от частоты импульсов ГнРГ[5]
Эпигенетическая регуляция функций гонадотрофов
гонадотропин
репродуктивный яичник гормон
В последнее время были достигнуты значительные успехи в понимании того,
как изменения структуры хроматина провоцируют угнетение транскрипции каких-либо
генов. Определённые трансформирующие ферменты обеспечивают посттрансляционную
модификацию «хвостов» гистонов, например ацетилирование или метилирование. Эти
модификации выражаются изменением генной экспрессии. Совсем недавно была
выявлена связь между модификациями хвостов гистонов и ключевыми процессами в
гипофизарных гонадотрофах, включая интересующие нас процессы транскрипции генов
β-субъединиц гонадотропинов и влияние
ГнРГ на эти процессы.
Ацетилирование и деацетилирование гистонов, ассоциированных с участками -
промоторами генов β-субъединиц гонадотропинов, играет важнейшую роль в
активации и супрессии этих генов. Этот динамический процесс связан с
вовлечением и активацией двух групп трансформирующих ферментов: гистоновых
ацетилтрансфераз (HATs) и гистоновых деацетилаз (HDACs). HATs ацетилируют
лизиновые остатки хвостов гистонов, и это рассматривается как процесс,
вызывающий локальную деконденсацию хроматина. Деконденсация, в свою очередь,
повышает доступность генных промоторных участков для транскрипционных факторов
и увеличивает возможность образования комплекса «РНК-полимераза - промотор» и
инициации транскрипции. Напротив, ферменты группы HDAC деацетилируют гистоны;
при этом их «хвосты» приобретают положительный заряд, и это приводит к возрастанию
компактизации хроматина (обусловленной электростатическими взаимодействиями
между отрицательно заряженной ДНК и положительно заряженными гистонами).
Некоторые из последних исследований приводят доказательства влияния ГнРГ на
стимуляцию генов β-ЛГ и β-ФСГ субъединиц, опосредованного
вышеописанной модификацией гистонов.
Исследования
более зрелых гонадотрофов LβT2-клеточной
линии, которые экспрессируют преимущественно мРНК βЛГ и, в меньшей степени, мРНК βФСГ, показали, что ферменты HAT опосредуют
ГнРГ-стимуляцию обоих видов β-субъединиц
гонадотропинов. HAT p300 ассоциирован с геном β-субъединицы ЛГ, и ГнРГ это связывание усиливает. В то
же время, p300, действуя совместно с транскрипционными факторами SF1, Sp1 и
Egr-1, играет важнейшую роль в усилении транскрипции β-субъедницы ЛГ. Было показано, что паралог p300 в LβT2-клетках,
CREB-связывающий белок (СВР), ассоциирован с промотором гена β-субъединицы ФСГ. ГнРГ может усиливать эту связь путём
взаимодействий с промотор-связанным фосфорилированным CREB, и стимулировать
таким образом транскрипцию β-субъединицы
ФСГ. (Рис.3) Дезактивация каким-либо образом р300 приводит к снижению транскрипции
обоих гонадотропных β-субъединиц, и в регуляцию транскрипции включается НАТ.
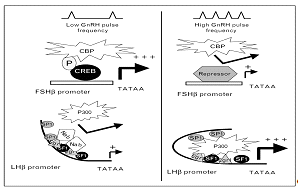
Рис.6. [5]
Итак, изучение αT3-1 и LβT2-клеточных линий подтверждает, что
модификация гистонов посредством динамического ГнРГ-контроля HDAC и HAT может
повлиять на присутствие/отсутствие ацетильных групп на хвостах гистонов,
ассоциированных с генами-регуляторами гонадотрофной транскрипции в целом и
транскрипции β-субъединиц гонадотропинов в частности. Так или иначе, нет
исследований, показывающих корреляцию между ассоциацией HDAC и HAT с
промоторами генов гонадотропинов и присутствием/отсутствием ацетильных групп на
гистонах. Будущие исследования, сосредоточенные в этой области, будут
способствовать дальнейшему пониманию относительной важности эпигенетических
модификаций в регуляции гонадотропинов, в особенности в ответ на пульсирующий
выброс ГнРГ. Кроме того, необходимо изучить другие модификации гистонов
(метилирование, фосфорилирование) с точки зрения регуляции ЛГ и ФСГ.
Заключение
ЛГ и ФСГ, синтез которых контролируется гипоталамическим декапептидом
ГнРГ, играют важнейшую роль в становлении и поддержании фертильной функции.
ГнРГ обладает пульсирующим характером секреции. Различные модификации
сигнальных каскадов, запускаемые ГнРГ в зависимости от частоты его выделения,
являются ключом к дифференциальному синтезу (и последующей секреции)
гонадотропинов. Изучение динамических характеристик и каскадов, запуск которых
инициирует ГнРГ, открывает огромные возможности для установления физиологических
механизмов овуляции и менструального цикла, таким образом способствуя развитию
новых виды терапии для лечения нарушений нейроэндокринного контроля
репродуктивной функции.
Список литературы
1Ciccone,
N. A. & Kaiser, U. B. The biology of gonadotroph regulation. National
Institute of Health 16, 321-327 (2009).
Гриффин,
Д. & Охеда, С. Физиология эндокринной системы. 158, 236-238 стр. (БИНОМ,
2008).
Armstrong,
S. P., Caunt, C. J., Fowkes, R. C., Tsaneva-Atanasova, K. & McArdle, C. A.
Pulsatile and sustained gonadotropin-releasing hormone (GnRH) receptor
signaling: does the ERK signaling pathway decode GnRH pulse frequency? J Biol
Chem 285, 24360-24371, doi:M110.115964 [pii]
.1074/jbc.M110.115964
[doi] (2010).
Caunt,
C. J., Finch, A. R., Sedgley, K. R. & McArdle, C. A. GnRH receptor
signalling to ERK: kinetics and compartmentalization. Trends Endocrinol Metab 17,
308-313, doi:S1043-2760(06)00150-0 [pii]
.1016/j.tem.2006.08.001
[doi] (2006).
Гриффин, Д. &
Охеда, С. Физиология эндокринной системы. Стр. 157 (БИНОМ, 2008).
1. Гриффин,
Д. & Охеда, С. Физиология эндокринной системы. Стр. 231 (БИНОМ, 2008).
. Гриффин,
Д. & Охеда, С. Физиология эндокринной системы. Стр.234 (БИНОМ, 2008).
3. Department
of biology and biochemistry. Research profile of J.Caunt, current research:
Mitogen activated
protein kinases and phosphatases in cell signaling (URL
<http://www.bath.ac.uk/bio-sci/research/profiles/caunt.html>)
4. Ciccone,
N. A. & Kaiser, U. B. The biology of gonadotroph regulation. National
Institute of Health 16, 321-327 (2009)