Генетика окрасов кошек британской породы
Генетика окрасов кошек
британской породы
Введение
Первое свидетельство об одомашнивании кошек, относится к 2500
году до н. э. и получено из Египта, где были обнаружены около сотни тысяч их
мумифицированных останков (Полладр, 2009 г.).
В Китае этих животных знали за тысячу лет до нашей эры, в
Японии о них узнали немного позже. Изображения кошек можно увидеть на рисунках
из Египта и Помпеи (Рис. 1). Египтяне обожествляли этих животных и считали, что
они обладают магическими свойствами.
К концу XVIII в Европе у кошек появилась репутация регулятора
численности грызунов. Это способствовало их распространению как необходимых
домашних животных (Коростовцев, 2008; Друбецкая, 2010).
Однако интерес к кошкам, как к экспонатам выставок,
пробудился лишь в конце XIX века. Люди стали отмечать различия в окраске их
шерсти и ее рисунках. А с зарождением и развитием генетической науки
специалисты по разведению кошек стали интересоваться созданием новых пород и
совершенствованием уже существующих (Друбецкая, 2010).
Кошки не могут быть традиционным объектом генетических
исследований. Они поздно становятся половозрелыми, плодовитость их невысока,
содержание этих животных в лабораторных условиях требует больших материальных
затрат. С другой стороны, широчайшая изменчивость собак и кошек и
"традиционность" их как физиологических объектов способствовали тому,
что целый ряд ученых посвятили себя изучению их генетики.
Исследования генома кошки важны и для человеческой медицины -
кошки имеют не менее 250 наследственных заболеваний, общих с человеком и
поражаются зачастую аналогичными инфекциями, что и человек. Домашняя кошка
предоставляет отличную модель для изучения инфекционных заболеваний, в том
числе модель для изучения человеческого СПИДа. (Бородин, 2010).
Начало исследований генетики кошки относится к 20-м годам XX
века (Бородин, 1993). Но некоторые данные о наследовании экстерьерных признаков
кошки обнаруживаются и в более ранних работах. Так, еще в XIX веке Чарльз
Дарвин анализировал происхождение домашней кошки и обращал внимание на огромное
многообразие ее форм. Он описывал бесхвостых кошек с острова Мэн, кошек с
висячими ушами из Китая, отмечал связь между белым окрасом шерсти, цветом глаз
и врожденной глухотой (Бородин, 2010).

Рис. 1. Наскальный рисунок кошки в Египте
В настоящее время проблемы генетики кошек привлекают все
большее и большее внимание ученых нашей страны. На сегодняшний день большинство
руководств по генетике кошек написаны специалистами по сельскохозяйственным
животным или молекулярной генетике, которые недостаточно хорошо ориентируются в
фелинологии в целом и часто слишком обобщенно судят о частной генетике животных
этих видов. Кроме того, часто статьи иностранных авторов, переводятся без учета
особенностей фелинологической терминологии, принятой у нас в стране. Иногда
представленные данные также не согласуются с принятой в России генетической
номенклатурой.
В то же время, несмотря на обилие фелинологической
литературы, практически нет руководств по генетике отдельных пород, описывающих
их специфику и селекционно-значимые признаки. Особенно это касается молодых
пород, к которым относятся и британские кошки. Использующие такую литературу
заводчики, по сути, являются любителями, а не профессионалами-селекционерами.
Незнание генетики своего объекта ведет к повышению уровня некомпетентности
Российских фелинологов.
Одним из важнейших селекционных признаков для кошек, является
их окрас. Знания по типированию - определению вероятного генотипа животного по
окраске и текстуре шерсти - позволяют проводить анализ наследования
селекционно-важных признаков у скрещивающихся британских короткошерстных и
длинношерстных домашних кошек.
Таким образом, на современном этапе развития фелинологии
большое значение приобретают точные методы определения окраса кошки. В
племенной работе с кошками часто встречаются случаи ошибочного определения
окраса при осмотре котят методистами племенных комиссий клубов любителей кошек
или выставочной оценке взрослых животных, что влечет за собой неадекватную
оценку генотипа и сомнения в генетической чистоте производителей. Наиболее
точное определение генотипа родителей позволяет правильно производить отбор и
подбор пар по такому важному в разведении кошек признаку, как окраска шерсти.
Встает вопрос о влиянии генов, определяющих окраску, длину и структуру шерсти,
на морфологию волоса не только на макро-, но и на микроскопическом уровне. Для
чего необходимо уточнить строение волоса кошки, поскольку имеющиеся в
литературе данные разрозненные и неполные.
Другая проблема, связана с тем, что большинство заводчиков не
рассматривают окрас шерсти кошки, как основной признак породы, но все же,
окраска шерсти для них селекционно-значима, в целях получения котят желаемого
окраса. Для скрещивания часто используют кошек с различными окрасами,
следовательно, кошки из питомника далеко не всегда имеют точно определенный
генотип относительно генов окраса.
Целью данной работы является изучение характера
наследования селекционо-важных окрасов у кошек британской породы на основе
литературных и полученных при анализе генотипов окрасов данных.
В соответствии с поставленной целью необходимо было решить
следующие задачи:
. Провести подробный анализ и сопоставление литературных
данных по генетике окрасов домашних кошек, и британских кошек в частности.
. Разработать методику и оценочные критерии, позволяющие
определять генотипы кошек британской породы по селекционно-важным генам
окрасов.
3. Определить генотипы по селекционно-важным окрасам
британских кошек питомника «Леонель» и британских кошек любителей.
. Выявить генотипы, формирующие классические окрасы (голубой
и лиловый) у кошек британской породы и оценить степень генотипической схожести
(гетерогенности или однородности) групп кошек одного фенотипа.
окрас кошка британский генетика
1. Обзор литературы
.1 Меланогенез у млекопитающих
Многообразие окрасок млекопитающих обусловлено наличием или
отсутствием пигментов меланинов. Меланины играют важную роль в клеточном
метаболизме, зрительной рецепции, обуславливают адаптацию кожных покровов к
внешней среде, защищая организм от действия ультрафиолетового излучения. Они
также присутствуют в мозговых оболочках и в меланоцитах опухолей. Поскольку
пигментные клетки закладываются в нервном валике на ранних стадиях
эмбриогенеза, то часто нарушения пигментации сопутствуют тяжелым поражениям
нервной системы и органов чувств (Сотская, 2010).
1.1.1 Формы меланина и распределение пигмента по
волосу кошки
Окрас шерсти зависит от типа пигмента, формы пигментных
гранул и распределения их по волосу. Меланины представлены двумя формами:
эумеланином и феомеланином. Меланины упакованы в пигментные зерна (меланосомы)
разнообразной формы. Пигмент может быть распределен равномерно или в виде
скопления гранул. Цвет волос зависит от вида пигмента, характера его
расположения и количества, а также содержания в волосе воздуха (Сотская, 2010).
По химическому составу эумеланины и феомеланины - полимеры
индол-5,6-хинона. Эумеланин присутствует в коже и волосах. Он нерастворим в
органических растворителях и устойчив к химической обработке. Феомеланин
образуется только в волосяных фолликулах и не встречается в коже. Эумеланин
имеет две модификации: черный − собственно эумеланин − и
коричневый, являющийся мутантной формой черного. Гранулы эумеланина придают
волосу высокую механическую прочность и упругость (Сотская, 2010). По
биофизическим характеристикам выделяют три модификации феомеланина: желтый,
оранжевый и красный. Структура клеток волос, содержащих только феомеланин,
характеризуется значительно меньшей прочностью, чем клеток, содержащих
эумеланин. Волосы, окрашенные феомеланином, менее устойчивы к механическим
воздействиям. Оба вида меланинов практически всегда присутствуют одновременно
(Сотская, 2010).
По волосу пигменты могут распределяться по-разному. У
животных окраса «агути», или зонарно-серого, эти пигменты, образуются в одной и
той же волосяной луковице и располагаются по длине волоса в определенной
последовательности (см раздел. 1.1.2). У кошек черного или коричневого окраса
феомеланин и эумеланин присутствуют в волосе. Но феомеланин «замаскирован»
эумеланином, поэтому волосы имеют черный или коричневый цвет (Сотская, 2010).
Плотность пигмента может быть неодинаковой в остевых,
покровных волосах и подшерстке. Пигментные гранулы, содержащие эумеланин, могут
разрушаться в клетках волоса. В этом случае меланин не равномерно распределен
по клетке. Пигментные гранулы могут быть равномерно распределены в клетках
различных слоев волоса, либо скапливаются в периферических или, наоборот, в
центральных отделах слоев. Темные остевые волосы содержат большие эллипсовидные
гранулы, а более светлые и тонкие волосы подшерстка - мелкие сферические.
При равномерном и плотном распределении пигмента тон окраски
интенсивный (Сотская, 2010).
Если интенсивно окрашена сердцевина волоса, а в корковом слое
пигмента мало или нет совсем, окрас выглядит осветленным (Сотская, 2010).
Многие окрасы меняются с возрастом животного. Это может быть
вызвано как изменением структуры шерстного покрова, так и перераспределением
пигментов в волосе, нарушением синтеза пигментов. Корковый и покровный слои
волос со временем частично стираются и мутнеют, поэтому волосы перед линькой
могут терять насыщенность цвета (Сотская, 2010).
1.1.2 Пигментогенез и формирование различных
окрасов у кошки
Палитру окрасов кошек составляют: черный, голубой,
шоколадный, коричневый, коричный, лиловый, бежевый, красный, кремовый и желтый
цвета. Белый цвет - это отсутствие пигмента (Шустрова, 1997). Пигментов у кошек
насчитывается относительно немного (см. раздел 1.1.1). Всё множество расцветок
окрасов кошек определяется генами окраски, влияющими на процесс формирования
окраса - пигментогенез (Шустрова, 1997).
Пигментогенез начинается в раннем эмбриогенезе. Закладка
меланобластов (будущих пигментных клеток) происходит у зародыша в области
нервной трубки (Шустрова, 1997). Дифференцировка меланобластов начинается с
изменения их формы. Они принимают веретенообразную форму, пригодную для
миграции. Они мигрируют сначала в центры пигментации (могут быть расположены в
виде пятен на темени, у корня хвоста, на спине или холке), а оттуда - в
волосяные фолликулы. Этот процесс находится под контролем гена White
(W) (ген доминантного белого окраса). Если этот ген представлен у
кошки двумя нормальными рецессивными аллелями w, клетки-предшественницы
приобретают нужную для миграции форму. Но если даже один из аллелей является
доминантным и мутантным W - клетки теряют способность к организации,
остаются на прежнем месте и в будущем пигмента производить не будут, а кошка -
носительница доминантного аллеля W останется неокрашенной (Шустрова,
1997).
Ген W может нарушать также формирование близлежащих
клеток радужки глаза и кортиева органа. В итоге образуются голубоглазые и даже
глухие кошки. В исследованиях Роя Робинсона было показано, что процент голубоглазых
кошек среди гетерозигот Ww незначительно ниже, чем среди гомозигот WW.
Также на экспрессивность гена W влияют гены модификаторы и регуляторные элементы
генома (Сингер, Берг, 1998). В настоящее время известно, что аллель W по
признаку собственно белого окрасам показывает почти полную экспрессивность и
100% пенетрантность, по признаку голубоглазости пенетрантность составляет около
40%, по глухоте - около 20% (Шустрова, 1997).
Для того чтобы меланобласты превратились в меланоциты, они
должны проникнуть в волосяной фолликул до его окончательного формирования (Рис.
2). Этот процесс определяет насколько окрашенной будет кошка и сохранятся ли у
нее белые пятна (Шустрова, 1997).
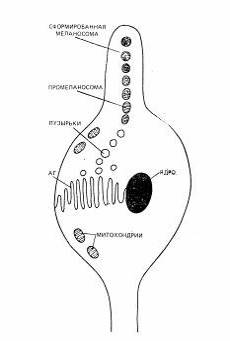
Рис. 2. Строение меланоцита и формирование меланосом
Равная роль в этом процессе отводится гену белой пятнистости
(пегости) - Spot, или, Piebald Spotting (S).
У кошек, гомозиготных по его рецессивному аллелю - ss - окраска
развивается полностью. А действие полудоминантного аллеля S вызывает ту
или иную степень белой пятнистости. Спектр пегости очень широк: от почти белого
окраса с цветным хвостом и пятнами на темени (ванский окрас) до почти полностью
развитого окраса с маленьким белым, «медальоном» на груди (Шустрова, 1997).
В последнее время часть фелинологов пришла к выводу, что у
гена белой пегости больше двух аллельных состояний (Шустрова, 1997).
Предполагают, что ванский окрас определяется наиболее доминантным аллелем серии
Sw, а арлекиновый окрас (цветные пятна разбросаны отдельными участками по
голове, холке, спине и крестцу) - вторым по степени доминантности аллелем Sp
(Шустрова, 1997). Отношения внутри пар этих аллелей не являются жестко
определенными, на них также сильно влияют гены-модификаторы (Шустрова, 1997).
Процесс превращения меланобластов в меланоциты контролируется
отдельной группой генов. Для превращения аминокислоты тирозина в промеланин, из
которого образуется меланин, необходим фермент тирозиназа (Рис. 10). А
образование этого регуляторного белка-фермента определяется геном, так называемого
локуса Colour (C). Этот локус представлен целой серией
аллелей. Доминантный аллель С обеспечивает синтез нормальной тирозиназы,
и тогда особь окрашивается полностью. Рецессивный аллель cs производит
термозависимую тирозиназу. Этот фермент активизируется только на холоде.
Поэтому гомозиготные по аллелю cs сиамские коты имеют интенсивную
окраску только наиболее охлаждаемых, выступающих участков тела - мордочки,
ушей, хвоста и лапок. Глаза у них голубые (Шустрова, 1997).
Сходная картина развивается при гомозиготности кошки по
другому аллелю серии - cb. Бурманский окрас сохраняет ту же
закономерность интенсивного окрашивания выступающих участков тела. При этом
глаза золотистые. Гетерозиготы cbcs - так называемые тонкинезы - имеют
промежуточный между сиамским и бурманским окрас и бирюзовый цвет глаз.
(Шустрова, 1997).
Другой ген - Black (B) - отвечает за синтез
меланина определенного цвета. При этом его доминантный аллель формирует
нормальную форму пигмента (черную), а рецессивный - окисленную (шоколадную).
Как и ген С, Black имеет аллельную серию - помимо аллелей черного
и шоколадного окраса, существует и наиболее рецессивный аллель bl,
образующий сильноокисленную форму эумеланина (Шустрова, 1997).
Сцепленная с полом, находящаяся на Х-хромосоме мутация Orange
(O), приводящая к развитию красных (рыжих) окрасов, приводит к
нарушению синтеза эумеланина, в результате чего клетки образуют только желтый
пигмент. У черепаховых кошек с генотипом Оо второй цвет зависит от
генетического набора аллелей в других локусах, и прежде всего в локусе В
(Шустрова, 1997).
При одном и том же количестве пигмента, окрас животного может
быть и ослабленным. Явление такого ослабления (осветления) окраса называется
разбавлением по Мальтесу. Определяется интенсивность окраса геном Dilutor
(символ D). Ген D отвечает за распределение гранул пигмента в
волосе. При наличии гена D гранулы меланина в волосе расположены плотно,
а аллеля d - рыхло. (Шустрова, 1997).
Некоторые фелинологи выделяют в локусе D еще один,
наиболее рецессивный аллель - dm. У кошек, гомозиготных по этому аллелю,
волоски ости осветлены и к кончику практически обесцвечиваются. В итоге кошка
голубого окраса как бы покрыта легким серебристым «напылением». Пример такого
окраса - шерсть русской голубой породы (Шустрова, 1997).
Лиловый цвет шерсти у кошек возникает в результате комбинации
рецессивных аллелей двух различных генов - мальтесианского осветления (его
генетический символ d) и шоколадного окраса (символ b). Окрасы
Павана (теплый золотисто-коричневый) и карамель (светлый золотисто-коричневый)
формируются под действием доминантного гена Dm - модификатора
осветления, действующего только при наличии в генотипе кошки
аллелей-разбавителей d (Шустрова, 1997). Павана формируется при участии
аллеля Dm на фоне голубого окраса, то есть при генотипе B-dd, а
карамель - на фоне лилового, при генотипе bbdd. Точные механизмы работы
этого гена пока что остаются невыясненными (Шустрова, 1997).
Отдельную группу представляют гены, определяющие характер
распределения пигмента по волосу кошки. Часто пигменты чередуются по длине
волоса зонами определенной толщины - это так называемый тикинг. Определяет
наличие тикинга доминантный ген локуса агути - Agouti (A), получивший свое
название от южноамериканского грызуна с таким окрасом (Шустрова, 1997).
Рецессивный аллель этого локуса носит название «неагути» (обозначается а)
и обеспечивает однотонный окрас волосков (Шустрова, 1997).

Рис. 3. Распределение пигмента по длине волоса: а - ровно
окрашенная; б - дымчатая; в - затушеванная
Обычно окрасы сочетаются с каким-то рисунком на теле кошки.
За наличие и тип такого рисунка отвечают аллели серии табби (Т - Tabby).
Очень редко, в естественных популяциях, встречаются кошки с ярко выраженным
тикингом, но практически без рисунка. Такой окрас характерен и для кошек
абиссинской породы (Шустрова, 1997).
Доминантный, аллель, отвечающий за формирование этого окраса,
называемого абиссинским или тикированным табби, обозначается как Та.
Гетерозиготы по этому гену, и редко гомозиготы - ТаТа имеют остаточные
элементы рисунка (кольца «ожерелья» на груди, слабые полосы на ногах и отметину
в форме буквы «М» на лбу). У кошек России наиболее распространен тигровый
рисунок (макрель-табби) - вертикальные полосы на туловище в сочетании с общими
для всех рисунчатых окрасов элементами (отметина «М» на лбу, кольца на груди,
завитки на щеках, два ряда двойных пятнышек вдоль живота и полосы на хвосте и
ногах). Этот рисунок определяется аллелем Т, рецессивным по отношению к Та.
Очень редкий в России, но обычный для Европы мраморный окрас кошек (включает
«бабочку» на плечах, две полосы вдоль спины и разводы на боках) характерен для
гомозигот по наиболее рецессивному аллелю серии табби - tbtb (Шустрова,
1997) (Рис. 3).
1.1.3 Строение и типы волос кошки
Макроскопическое строение волоса кошки
Макроскопически различаются следующие части волоса: луковица,
или корень волоса (1), шейка луковицы (2), тело волоса (3), кончик волоса -
типпинг (4) (Рис. 4а) (Голубева, 2008).
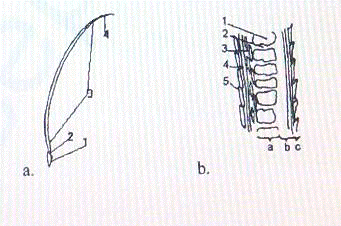
Рис. 4. Схема макро - (а) и микростроения (b) волоса кошки
В поперечном сечении волос разделяется на 3 слоя:
сердцевинный или медула (а), корковый (b) и чешуйчатый (с). При
микроскопии можно выделить следующие структуры: полости сердцевинного слоя
заполненные воздухом (1), белковые трабекулы (2), веретенообразные клетки
коркового слоя (3), субкутикулярная мембрана (4), чешуйчатый слой (5) (Рис. 4b).
Таблица 1. Типы волос кошки

По морфологическому строению выделяют четыре основных типа
волос кошки (Таблица 1).
1) Вибрисы (чувствительные), располагаются над верхней
губой, над глазами, на щеках и на плантарной поверхности пясти на
чувствительных тельцах. Корень вибрисы толстый и длинный, в нижней части имеет
углубление, для нервного окончания. Шейка не выражена, тело волоса толстое.
Сердцевидный слой отсутствует. В теле вибрисы обнаруживаются небольшие полости
вытянутые продольно (Голубева, 2008).
2) Покровные волосы хорошо развиты. Их количество
зависит от породы кошки и от расположения на теле. Больше покровных волос на
спине и на хвосте, меньше на животе. Они составляют украшающий волос на щеках,
на воротнике, на очесах передних и задних лап. Корень покровного волоса имеет
грибовидную вытянутую форму. Шейка хорошо выражена, лишена пигмента. Средний
слой или медула начинается веретенообразно в месте выхода волоса из кожи кошки.
Она представляет собой цилиндрическую полость с фестончатым краем. Поперек
медулы лежат трабекулы, состоящие почти целиком из кератина. Между трабекулами,
в результате разрыва клеток, образуются полости, заполненные воздухом. Форма
трабекул и полостей, определяемая аллельным состоянием генов окраски может быть
ровной или деформированной. Медула заканчивается, немного не доходя до кончика
волоса. Снаружи от медулы лежит плотный корковый слой, если кошка окрашена, то
веретенообразные клетки коркового слоя содержат пигмент, соответствующий
аллелям, находящимся у нее в генотипе. Кутикула представляет наиболее поверхностный
слой волоса. Она состоит из ороговевших чешуек. Они накладываются друг на друга
наподобие черепиц. Их свободные концы направлены по направлению от корня волоса
к кончику. На здоровом покровном волосе они плотно прилегают и лишь слегка
заметны. Кутикула играет большую роль в формировании тона и интенсивности
окраса, а также блеска шерсти. Кончик покровного волоса состоит из коркового
слоя и чешуек кутикулы. При наличии пигмента в черном или типированном волосе
кончик окрашен особенно интенсивно (Голубева, 2008).
) Пуховые волосы (подпушь) составляют подшерсток,
густота которого зависит от породы кошки. Подпушь растет розетками по несколько
волосков около одного покровного. Морфологически пушковые волосы разнообразны
даже у одного животного. Они различаются по длине, толщине и степени
выраженности медулы. Их объединяют некоторые черты: слабый корень овальной
формы, короткая шейка, более широкие и рыхло расположенные трабекулы, сильная
зазубренность чешуек. При наличии окраски, пушковые волосы содержат меньше
пигмента и в целом выглядят светлее, чем покровные.
) Пух - самые тонкие волоски. Состоит только из
чешуек. Средний слой отсутствует. Корень очень слабый, округлой формы. Пух
обнаруживается у пород европейского происхождения. Поскольку пух тоньше и не имеет
медулы, его окраска всегда слабее, чем у других типов волос (Голубева, 2008).
Микроскопическое строение волоса кошки
Гены окраски в различном аллельном состоянии влияют на
микроскопическую структуру волоса (Робинсон, 1995). Это характерно для всех
типов волос, но наиболее выражено в направляющих и остевых волосах (Робинсон,
1995).
Белый волос, сформировавшийся под влиянием доминантных
аллелей локусов Dominant White, Piebald Spotting и Melanin Ingibitor (нижняя часть волоса)
имеет прозрачный корковый слой, лишенный пигмента (Рис. 5) (Голубева, 2008).

Рис. 5. Микрофотография белого покровного волоса (W-) (увеличение 1х400)
Черный пигмент (эумеланин) плотно заполняет веретенообразные
клетки коркового слоя. Особенно плотно пигмент упакован на кончике волоса
(типпинг), поскольку там отсутствует сердцевина. Далее к сердцевине волоса и,
особенно у корня, наблюдается более рыхлое расположение трабекул в сердцевинном
слое и гранул пигмента в корковом слое.
У черной кошки типпинг волоса выглядит интенсивно черным (Рис
6). У корней волоски приобретают коричневатое или серо-голубое окрашивание.

Рис. 6. Микрофотография черной вибрисы (аа)
Под влиянием аллеля Тa происходит так называемое
«осветление эумеланина по абессинскому типу», захватывающее нижнюю часть
волоса. Черный пигмент приобретает рыжевато-коричневую окраску (Голубева,
2008).
Аллели brown и brown light вызывают окисление
эумеланина до коричневого и коричного. У кошки, несущей одну из этих аллелей в
гомозиготе типпинг на всех волосках полупрозрачного коричневого оттенка, при
компактном расположении пигмента (Голубева, 2008).
Рецессивный аллель d локуса Dilution вызывает слипание
пигмента в гранулы, деформацию трабекул и полостей медулы. Эти гранулы хорошо
различимы при микроскопии (Рис. 7) (Голубева, 2008).
Состояние пигмента внутри гранул зависит от наличия других
аллелей в генотипе кошки. За счет появления пустот в стержне волоса визуально
окрас осветляется. В спорных случаях аллель d определяют по типпинговой
части волоса, где кутикула становится прозрачной. Деформацию дисков медулы
способен вызвать доминантный аллель I, что хорошо заметно в
нижней осветленной части волоса. При этом пигмент в типпинговой части компактно
заполняет корковый слой. Под влиянием генотипа dd bb и dd bibi формируются окрасы
лиловый и фавн. Корковый слой волоса у таких кошек прозрачный, содержит
отдельные гранулы осветленного пигмента (Рис. 8) (Голубева, 2008).

Рис. 7. Микроскопия осветленного покровного волоса (dd) (увеличение 1х400)

Рис. 8. Микрофотография лилового волоса подпуши (dd bb) (увеличение 1х400)
При тикированных окрасах (А-) в волосе присутствуют оба
пигмента. Эумеланин всегда заполняет верхушку волоса и распределяется по телу
волоса. Феомеланин формирует кольца тикинга (от 1 до 18). Красный пигмент
феомеланин при компактном расположении в кутикуле выглядит оранжевым, а в
дисках медулы коричневым. Красный волос при генотипах ОО / Оу Оо (локус Оrange)
и красная зона тикинга в волосе агути всегда выглядит более прозрачной, чем
волос, окрашенный феомеланином (рис. 9) (Голубева, 2008).

Рис. 9. Микрофотография покровного волоса окраса агути (А-)
в месте перехода колец тикинга
На участках окрашенных феомеланином просматривается
продольная исчерченность коркового слоя, обусловленная веретенообразными
клетками. Эта группа окрасов трудно дифференцируется от коричневого (bb и bibi). Абиссинский красный (Ta-) отличается от красного
сцепленного с полом (Orange) наличием интенсивного типпинга (Голубева, 2008).
.2 Генетика окрасов кошек
Окраска определяется функционированием взаимодействующих
генов, проявляющих активность в различные периоды онтогенеза и в разных тканях
или действующих во всем организме в одно и то же время (Ватти, Алексеевич,
1976).
Таблица 2. Основные группы генов окрасок у кошек
|
Гены окрасок
шерсти у кошек
|
Обозначение
|
Аллельные
варианты
|
Фенотипический
эффект
|
|
1. Гены,
определяющие характер распределения пигмента по длине волоса
|
|
|
Серия агути - А
- Agouty (А > а)
|
А
|
Агути -
зонарный окрас каждого волоска
|
Волосы
короткошерстных кошек окрашены равномерно от основания волоса до кончика. У
длинношерстных кошек наблюдается постепенное снижение интенсивности окраски к
основанию волоса. Для кошек «агути» окраса характерно наличие светлой
отметины в форме отпечатка большого пальца человеческой руки на задней
поверхности уха, а также розовой или кирпично-красной мочки носа, окаймленной
узкой темной полосой
|
|
а
|
Не агути
-сплошной окрас каждого волоска по всей длине
|
|
|
2.
Гены, определяющие цвет пигмента
|
|
|
Серия черного
цвета - B - Black. Отвечает за синтез эумеланина и
соответственно за проявление черного / коричневого окраса (B > b > b l)
|
B
|
Не окисленный
эумеланин
|
Черный. Нет
шоколадный окрас волосков
|
|
b
|
Окисленная
форма пигмента эумеланина - феомеланин
|
Шоколадный
окрас
|
|
b l
|
Еще более
коричневый окрас
|
Цвет корицы,
циамона. Породы, у которых встречается данный окрас: сомалийская,
абиссинская, австралийский микст
|
|
Серия красных
окрасов (нет образования черного пигмета эумеланина, а только желтого
феомеланина - O - Orange (сцеплены с Х-хромосомой) (О > o)
|
OO
|
Красная кошка
(или ее осветленные формы)
|
|
|
Oy
|
Красный кот
(или его осветленные формы)
|
|
|
OX (или Oo)
|
Черепаховая
кошка (или ее осветленные формы)
|
|
|
XX (или oo)
|
Не красная
кошка (нет гена О)
|
|
|
XY (или oy)
|
Не красный кот
(нет гена О)
|
|
|
3. Гены,
определяющие степень окрашенности
|
|
|
Серия
альбиностических аллелей - С - Color (C > c b > c s > c a
> c)
|
C
|
Полное
окрашивание всего тела кошки
|
Сплошные окрасы
|
|
c b
|
Пойнты
(отметины)
|
Лапы, хвост,
морда, уши темнее общего фона тела (бирманский окрас), осветление цвета глаз
до золотистого
|
|
c s
|
Светлое тело с
темными пойнтами (отметинами)
|
Окрас корпуса
значительно светлее окраски мордочки, лап и хвоста - цвета топленого молока и
светлее. Для таких кошек типичен голубой окрас радужной оболочки (колор-пойнт
или сиамский альбинос). Между аллелями cs и cb наблюдается
промежуточное доминирование. Гетерозиготы cbcs (тонкинезы) имеют глаза
бирюзового цвета и окрас, похожий на сиамский и бурманский одновременно
|
|
c a
|
Полный
альбинизм
|
Белый окрас
шерсти, светло-голубой цвет глаз с полупрозрачными зрачками
|
|
c
|
Полный
|
Белая окраска
шерсти, радужная оболочка прозрачна и светится розовым (за счет
просвечивающихся капилляров), зрачок также розовый. Встречается чрезвычайно
редко
|
|
4. Гены,
определяющие интенсивность окраски
|
|
|
Серия
интенсивности окрашивания волосков (осветления) - D - Dilition, Dense
pigmentation. (D > d
> d l)
|
D
|
Волос окрашен
интенсивно
|
Базовый - не
осветленный цвет: черный, красный, шоколадный
|
|
d
|
Волос осветлен
|
Голубой,
кремовый, лиловый. Распределение пигментных гранул в радужной оболочке глаз
нормальное.
|
|
d l
|
Только кончик
осветленного волоска не имеет пигмента (dilution light)
|
Окрас русских
голубых кошек
|
|
Серия
серебристости (дымов) (неокрашивание нижней части волосков у основания) - I
- Melanin Inhibition (I > i)
|
I
|
Неокрашивание
нижней части волоса (дымы, серебристые)
|
|
|
i
|
Нормальная
прокраска каждого волоска
|
Отсутствие
серебристости (нормальный, полный окрас)
|
|
5. Гены,
определяющие распределение пигмента по телу и пятнистость
|
|
|
Серия белой
пегости (биколоры, медальоны) - S - Piebald Spotting (S v> S p > S > s > s i)
|
S
|
Есть белые пятна
|
|
|
s
|
Белых пятен нет
|
|
|
S v
|
Ванский окрас
(не менее 1/3 белого)
|
|
|
S p
|
Арлекиновый
окрас (не менее 1/2 белого)
|
|
|
S
|
Биколорный
окрас (не более 1/2 белого)
|
|
|
s
|
Окрас без
белого
|
|
|
Серия белых
«перчаток» (лапок) (рэгдоллы) - Gl - Gloves (Gl > gl)
|
Gl
|
Белых
«перчаток» нет
|
|
|
gl
|
Белые
«перчатки» есть
|
|
|
Серия окрасов с
рисунками - T - Tabby (T a > T > t b > t s)
|
T a
|
Сплошной.
|
Дикий
абиссинский окрас. Полосы только на морде, лапах и хвосте, но каждый волос
зонарно окрашен полосами разного цвета
|
|
T
|
Тигровый
|
Полосатый.
Рисунок окраса кошки
|
|
t b
|
Мраморный
рисунок
|
Замкнутые
кольца-бабочки
|
|
t s
|
Пятнистый
рисунок окраса кошки
|
|
|
Серия
доминантного сплошного белого окраса - W - Dominant White (W > w)
|
W
|
Полностью белая
кошка
|
|
|
w
|
Не белая
(цветная) кошка
|
|
1.2.1 Комбинации генов, определяющие разные типы
окрасов
Комбинации генов, определяющие черный и голубой окрасы
Черный окрас кошек определяет рецессивная пара aa и
доминантные аллели В-, С-, D- (Сотская, Московкина, 2000) (Рис. 11). У
короткошерстных кошек подшерсток часто окрашен также ярко, как покровные
волосы. У длинношерстных в большинстве случаев подшерсток коричневый или
сероватый (Сотская, Московкина, 2000)
При взаимодействии пары рецессивных генов dd черный
окрас осветляется и превращается в голубой ааB-C-dd (Сотская,
Московкина, 2000г) (Рис. 12).
Под воздействием гена-ингибитора I происходит
обесцвечивание основания волос длинношерстных кошек, что приводит к образованию
дымчатых окрасов (Сотская, Московкина, 2000) (Рис. 13).

Рис. 11. Кошка черного окраса

Рис. 12. Кошка голубого окраса

Рис. 13. Кошка черного дымчатого окраса
Комбинации генов, определяющие коричневые окрасы
Коричневый окрас определяется генами аа и bb и
доминантными С-, D. Кожные покровы, мочка носа и подушечки лап
таких кошек также имеют коричневую окраску (Сотская, Московкина, 2000)
Интенсивность коричневого окраса может определяться другими
аллелями локуса b, а так же наличием аллели dd:
bbDD - шоколадный окрас (Рис. 14)
b, b, DD - светло - коричневый (циамон) (Рис. 15).
bbdd и b, b, dd - лиловые (Рис. 16) или сиреневые.
Воздействие гена-ингибитора I на окрасы этой группы
приводит к образованию шоколадных или лиловых дымчатых окрасов (Сотская,
Московкина, 2000 г.)

Рис. 14. Кошка шоколадного окраса

Рис. 15. Кошка светло-коричневого окраса (циамон)

Рис. 16. Кошка лилового окраса
Комбинации генов, определяющие белый окрас
Белый окрас (Рис. 17) кошек в большинстве случаев обусловлен
доминантным аллелем W. Иногда доминантным аллелем S, а так же
альбиностическими аллелями сс и саса (Сотская, Московкина, 2000).

Рис. 17. Кошка белого окраса
Комбинации генов, определяющие рыжий окрас
Рыжий (красный) окрас (Рис. 18) определяется преимущественно
доминантным геном О, расположенным в половой хромосоме. Возможно, у
кошек имеется несколько типов феомеланина, которые могут определять разные тона
рыжего окраса. Кошки - носители аллелей T или tb - полосатые,
пятнистые или мраморные. Носители Ta могут быть однотонно рыжими
(Сотская, Московкина, 2000).
Аллели dd вызывают ослабление красного и рыжего окраса
до кремового.
Ген-ингибитор I у длинношерстных кошек способствует
формированию окрасов группы камео. Кожные покровы при рыжих окрасах кошек не
пигментированные. Рыжий окрас могут иметь и абиссинские кошки, носители аллелей
bb (Сотская, Московкина, 2000).

Рис. 18. Кошка рыжего окраса
Комбинации генов, определяющих сиамские окрасы
Аллели сиамского альбинизма сscs в сочетании с другими
аллелями, влияющими на расцветку шерсти, определяют целую гамму окрасок.
Основной окрас в соответствии с формулой проявляется на морде, ногах и хвосте.
Корпус, шея и верхние части ног у кошки осветлены.
Таблица 4. Генотипы сиамских окрасов
|
aaB-cscsD
-
|
Seal-point
|
Отметины
интенсивного темно-коричневого окраса, тело светло-бежевое, мочка носа и
подушечки лап почти черные.
|
|
aaB-cscsdd
|
Blue-point
|
Отметины
голубовато-серого окраса. Тело светлое с голубоватым оттенком, Мочка носа и
подушечки лап серые.
|
|
aabbcscs
D -
|
Chocklate-point
|
Отметины цвета
молочного шоколада. Тело цвета слоновой кости. Мочка носа и подушечки лап
коричневые.
|
|
aabbcsc
sdd
|
Lilac-point
|
Отметины
лилового окраса, тело практически белое. Мочка носа и подушечки лап розовые с
легким сиреневым оттенком.
|
|
csc
sD - OO
|
Red-point
|
Отметины
оранжевого, рыжего или красного цветов, тело с легким кремовым оттенком.
Мочки носа и подушечки лап розовые.
|
|
aa csc
sddOO
|
Cream-point
|
Отметины
кремового цвета. Тело с легким, кремовым оттенком. Мочка носа и подушечки лап
розовые.
|
|
aaB
- csc s D - Oo
|
Seal-tortie-point
|
С
темно-коричневыми черепаховыми отметинами.
|
|
aaB
- csc s dd Oo
|
Blue-tortie-point
|
С голубыми
черепаховыми отметинами.
|
|
aabb
csc s dd Oo
|
Lilac-tortie-point
|
С лиловыми
черепаховыми отметинами.
|
|
aabbcsc
s D - Oo
|
Chocolate-tortie-point
|
С шоколадными
черепаховыми отметинами.
|
|
A -
B - cscsD - T - (tbtb)
|
Seal-tabby-point
|
C темно-коричневыми полосатыми
отметинами.
|
|
A -
B - cscsddT - (tbtb)
|
Blue-tabby-point
|
С голубыми
полосатыми отметинами.
|
|
A -
bbcscsD - T - (tbtb)
|
Chocolate-tabby-point
|
С шоколадными
полосатыми отметинами.
|
|
A -
bbcscsddT - (tbtb)
|
Lilac-tabby-point
|
С лиловыми
полосатыми отметинами.
|
|
A -cscsD
-OO T - (tbtb)
|
Red-tabby-point
|
С красными
полосатыми отметинами.
|
|
A -
cscsddOO T - (tbtb)
|
Cream-tabby-point
|
С кремовыми
полосатыми отметинами.
|
Кошки гетерозиготные по аллелю О(Оо) имеют отметины
черепахового окраса в соответствии с основной формулой окраса (Сотская,
Московкина, 2000).
Сиамские окрасы на фоне полосатых окрасов имеют полосатые
отметины и называются тебби-пойнт (Сотская, Московкина, 2000).
Бурманские окрасы определяются генами cbcb (Сотская,
Московкина, 2000).
Генотипы бурманских окрасов:
aaB- cbcbD- - коричневая бурма,
aaB- cbcbdd - голубая бурма,
aabb cbcbD- - шоколадная бурма,
aabbcbcbdd - лиловая бурма,
aа cbcbD-ОО - красная бурма,
aacbcbddOO - кремовая бурма.
Носители аллелей cbcb и Оо представляют собой
группу черепаховая бурма, и т.д (Сотская, Московкина, 2000).
Комбинации генов, определяющие абиссинский окрас
Абиссинский (Рис. 19), или сомалийский, окрас определяется
аллелем Tb. Кошка имеет равномерный без полос окрас. Волосы имеют
зонарное окрашивание с двойным или тройным тикингом с чередованием
красно-коричневого и черного цвета. Кончики остевых волос окрашены в черный
цвет. Волосы подшерстка, а также кроющие волосы на животе, груди и внутренней
стороне ног не имеют тикинга. Поэтому эти участки тела имеют ровный рыжий окрас
различных оттенков (Сотская, Московкина, 2000) (Таблица 5).
Таблица 5. Генотипы абессинских окрасов
|
A-B-C-D-Ta
|
Дикий окрас
абиссинского типа.
|
|
A-bbC-D-Ta
|
Красно-коричневый
абиссинский (соррель) окрас. Варьирует от рыжевато-коричневого до
медно-красного цвета. Подшерсток и основания остевых волос окрашены в более
темный цвет. В данном случае красный окрас кошки обусловлен не сцепленным с
полом аллелем О, а тем, что черные зоны волоса заменены на коричневые,
поэтому волос выглядит почти равномерно красным. Мочка носа кирпично-красная
с красно-коричневым ободком. Подушечки лап розовые.
|
|
A-B-C-ddTa
|
Голубой
абиссинский. Зонарность волоса обусловлена чередованием голубых и
светло-кремовых полос, что придает стально-глубокий оттенок всей шерсти
кошки. Мочка носа кирпично-красная с голубой окантовкой. Подушечки лап
серо-голубые.
|
|
A-bbC-ddTa
|
Бежевый (цвет
косули). Зонарность волоса представлена темными и светло - кремовыми зонами.
Мочка носа розовая с грязновато-розовой окантовкой. Подушечки лап также
грязно-розовые.
|

Рис. 19. Кошка абиссинского окраса
1.2.2 Гены, определяющие распределение пигмента
по телу кошки
Окрасы с неравномерным распределением пигмента по поверхности
тела чаще всего обусловлены действием аллеля I (Melanin Ingibitor)
на фоне длинной шерсти. Также пятнистость вызывают различные алельные сочетания
генов S и O. Кроме того, при наличии в генотипе агути-фактора
кроме зонарного окраса волоса проявляется тот или иной тип рисунка на корпусе.
Пегий окрас является проявлением фактора белой пятнистости. Агути - явление
зонированного окрашивания волоса. К группе агути-окрасов у кошек относятся
окрасы табби, затушеванные, «шиншиллы».
Комбинации генов, определяющие дымчатые окрасы
Эта группа окрасов образована взаимодействием аллелей aa
и I-. При этом остевые волосы обесцвечены от корня так, что окрашенной
остается 1/3 часть верхушечной части волоса. Подшерсток практически белый
(Сотская, Московкина, 2000). Генотипы дымчатых окрасов представлены в таблице
6.
Таблица 6. Генотипы дымчатых окрасов
|
aaB-D-I-
|
Черный дымчатый
|
|
aabbddI-
|
Голубой
дымчатый
|
|
aabbD-I-
|
Шоколадный
дымчатый
|
|
aabbddii
|
Лиловый
дымчатый
|
Комбинации генов, определяющие окрасы камео
Группа окрасов обусловлена взаимодействием аллелей I-
и OO. В группе выделяют: красный камео, красный затушеванный камео и
красный дымчатый. Все эти окрасы имеют одинаковый генотип D-I-OO,
различия, вызваны степенью экспрессии гена I и действием генов-модификаторов.
Отличия между ними заключаются в длине окрашенной части остевых волос. У камео
окрашенная часть составляет 1/8 волоса, у затушеванного камео - 1/3 волоса, у
дымчатого - остевые волосы окрашены почти до самого корня. Подшерсток у всех
камео белый (Сотская, Московкина, 2000) (Рис. 20).

Рис. 20. Кошка окраса камео
Комбинации генов, определяющие шиншилловые окрасы
Шиншилловый окрас - это светлые волосы с темными концами. К
этой же группе относятся и дымчатые окрасы. Эти окрасы наблюдаются в основном у
длинношерстных кошек (Сотская, Московкина, 2000).
Принято выделять 2 типа шиншилловых кошек: серебристые (Рис.
21) и золотистые (Рис. 22). У серебристых шиншилл подшерсток и основания
остевых волос имеют золотисто-рыжий или золотисто-желтый цвет (Сотская,
Московкина, 2000).
В зависимости от степени прокрашенности (тикинга и типпинга)
отдельных волос и их оттенков, как серебристые, так и золотистые шиншиллы имеют
целый ряд вариантов окраса (Сотская, Московкина, 2000).
Золотой оттенок зависит от генов руфизма (обеспечивают
дополнительный синтез феомеланина) (Робинсон, 1995).
Таблица 7. Взаимодействие аллелей серебристого и золотого
окрасов
|
Аллели
|
Эффекты на
генетическом фоне
|
|
Bleacher
|
Eraser
|
Агути (А-)
|
Неагути (аа)
|
|
Bl Bl
|
Er Er
|
серебристая
шиншила
|
дымчатый окрас
|
|
Bl Bl
|
Er er
|
серебристая
затушёванная
|
дымчатый окрас
|
|
Bl Bl
|
er er
|
серебристая
табби
|
слабый дым
|
|
Bl bl
|
Er Er
|
серебристая
шиншила или затушёванная
|
дымчатый окрас
|
|
Bl bl
|
серебристая
затушёванная или табби с желтизной
|
дымчатый окрас
|
|
Bl bl
|
er er
|
серебристая
тебби
|
слабый дым
|

Рис. 21. Кошка окраса - серебристая шиншилла

Рис. 22. Кошка окраса - золотистая шиншилла
В настоящее время наиболее распространены «бигенные» теории
золотых и серебристых окрасов (базируются на двух отдельных локусах)
(Шустрова,1997).
Предполагается, что гены, отвечающие за серебристый окрас
(ингибиторы меланина и феомеланина) действуют независимо от генов золотого
окраса - ингибиторов эумеланина, черного. Эти гены были недавно названы Bleacher
и Eraser соответственно (название и генетические символы неофициальные).
Каждый из этих генов должен быть представлен как минимум двумя аллелями,
действующими, естественно, на агути или неагути-фоне (Шустрова,1997).
Комбинации генов, определяющие пятнистость
Пятнистый окрас (spotted), состоящий из приблизительно
ровных, одинаковых по размеру пятнышек на боках - один из самых сложных
вариантов окраса у кошек. При скрещивании между собой пятнистые кошки всегда
дают котят только с таким же рисунком. Неизвестны и случаи появления пятнистых
котят у пар мраморного окраса. Между тигровым и пятнистым окрасом наблюдаются
все переходные степени - от разорванных в нескольких местах полос до почти
круглых пятен. Известны и такие особи, у которых 2-3 полосы на передней части
тела сочетаются с пятнами на задней. Это могло бы быть объяснено промежуточным
проявлением аллелей Т и tsp. На сотни кошек с промежуточными
тигрово-пятнистыми окрасами известны лишь единицы с разорванным мраморным
рисунком.
Существует гипотеза, что табби (Рис. 23) представляет собой
сложный, протяженный локус, внутри которого возможен обмен участками -
генетическим материалом (внутригенная рекомбинация). Согласно другой гипотезе,
механизм развития рисунка связан с внутренними биологическими часами развития
организма. У маленьких белых котят можно заметить такой «рисунок», образованный
полосками волос разной длины и текстуры: более длинные и грубые чередуются с
более тонкими и короткими (Шустрова,1997).
Комбинации генов, определяющие черепаховый окрас
Черепаховая окраска может быть только у особей с двумя Х-хромосомами.
Расположение пятен зависит от случайного характера инактивации Х-хромосомы.
Цвет не рыжих пятен зависит от аллелей разных локусов, определяющих окрас. Так,
при генотипе aaB-C-D- - пятна черные, при генотипе A-B-C-D-T- -
пятна полосатые и т. д (Шустрова,1997).
При полосатых окрасах рыжие пятна четко вписываются между
полосами рисунка, создавая впечатление практически рыжего фона. Но иногда
напротив, рыжие пятна бывают столь малы и редки, что могут быть почти не
заметны, особенно на фоне общего ослабления окраски и у длинношерстных кошек
(Сотская, Московкина, 2000).
Генотипы черепаховых кошек:
A-B-C-D-Oo-T- - рыжие и черно
полосатые пятна (Рис. 23);
A-B-C-D-Ootbtb - - рыжие и черно-мраморные
пятна (Рис. 24);
При этом расположение полос одинаково на черном и рыжем фоне;
A-B-C-dd-Oo-T- и A-B-C-dd-Oo tbtb - то же самое, только
ослабленного окраса;
AaB-C-D-Oo - черные и рыжие пятна
без полос (Рис. 25);
AaB-C-dd-Oo - голубые и кремовые пятна
(голубой черепаховый или голубо-кремовый окрас) (Сотская, Московкина, 2000 г.)
(Рис. 26).
У котов черепаховый окрас (Рис. 25) может присутствовать,
только если кроме Х- и Y- хромосом, он имеет еще одну лишнюю Х-
хромосому, и его хромосомная конституция будет обозначена, как ХХY
(аналогично синдрому Кляйнфельтера у человека). Но недавно, были обнаружены
черепаховые коты, имеющие одну Х-хромосому (Бородин, 2009).

Рис. 23. Кошка окраса - полосатый тебби

Рис. 24. Кошка окраса - пятнистый тебби

Рис. 25. Кошка черепахового окраса

Рис. 26. Кошка голубо-кремового окраса
1.2.3 Гены, влияющие на окрас кожных покровов у
кошек-сфинксов
Сейчас существуют три породы голых кошек-сфинксов: канадский
сфинкс и две - выведенные в России - донской сфинкс (лысак) и петерболд
(Нестерова, 2007).
Белый: цвет кожи от белого до розового. Кожа носа и подушечки
лап - розовые.
Черный: один тон черного окраса от носа до кончика хвоста.
Зеркальце носа черное. Подушечки лап черные или коричневые.
Голубой: один тон окраса от носа до кончика хвоста. Зеркальце
носа и подушечки лап голубые.
Красный: глубокий, насыщенный, ясный, красный цвет без
маркировок. Губы и подбородок цвета, аналогичного цвету кожи. Зеркальце носа и
подушечки лап красно-коричневые.
Кремовый: один оттенок кремового по всему телу, без
маркировок. Зеркальце носа и подушечки лап розовые.
Шоколадный: насыщенный красновато-коричневый, распределение
по телу ровное. Зеркальце носа коричневое. Подушечки лап коричневые или цвета
корицы.
Лиловый: морозно-серый с розоватым оттенком, ровного
распределения. Зеркальце носа и подушечки лап розовые, лавандовые.
Циамон: цвет корицы, равномерно распределенный по всему телу.
Зеркальце носа и подушечки лап цвета корицы.
Фавн: бледный светло-бежевый цвет равномерно распределенный
по всему телу. Зеркальце носа и подушечки лап бледно-бежево-розовые (Нестерова,
2007).
.2.4 Гены, влияющие на длину и текстуру шерсти
Длина шерсти определяется аллельным состоянием гена Long
(L). Рецессивный аллель этого гена удлиняет время роста волоса, за
счет чего у гомозигот ll развивается удлиненная шерсть. Однако
проявление этого гена отличается большим разнообразием, поддающимся
направленной селекции (Шустрова,1997).
Изменение текстуры шерсти определяют рекс - мутации. У
рекс-мутантов волоски извитые с нарушенным строением. Вибриссы также извитые.
Ген рексовости наследуется по рецессивному типу и имеет обозначение r.
На основе мутации корниш-рекс создана отдельная порода под тем же названием.
В Германии обнаружена рекс-мутация, давшая начало породе
немецкий рекс. Эта мутация оказалась аллельна к r. Генотип немецкого
рекса- rgrg (Шустрова, 1997).
Другая мутация - проволочная шерсть - характеризуется
доминантным типом наследования (Wh - Wire hair). В ее основе
лежит американская жесткошерстная порода кошек. Покровный волос и ость тоньше,
чем у обычных кошек. Остевые волосы очень изогнуты, почти надломаны. А волоски
подшерстка утолщены и так же изогнуты. На ощупь шерсть грубая и упругая
(Шустрова, 1997).
Мутация «ватная» шерсть (самсоновость) определяется
рецессивным аллелем sa (Satin). Шерсть гомозигот по этому аллелю
тонкая и густая строение волос нарушено, следовательно, они теряют жесткость.
Их покровную шерсть, из-за нарушения структуры, волос трудно отличить от
остевой (Шустрова, 1997).
Отсутствие шерсти (атрихоз) - явление, представленное
представлено целой группой неаллельных мутаций. В настоящее время изучено
наследование только трех вариантов бесшерстности кошек (Шустрова, 1997).
Французская бесшерстность - мутация с рецессивным
наследованием (h). Котята рождаются покрытыми детской пуховой шерсткой,
которая впоследствии исчезает. Иногда у взрослых животных сохраняются настоящие
волосы, особенно на ногах, хвосте и мордочке. Вибрисы прямые (Шустрова, 1997).
Канадская бесшерстность. Внешне схожа с французской, но
относится к другому локусу - hr - и также определяется рецессивным
геном. На ее основе создана порода канадский сфинкс (Шустрова, 1997).
Мутация hd. У гомозигот по этой мутации кожа при
рождении мягкая и эластичная, но с возрастом становится толстой и собирается в
складки (Шустрова, 1997). Вибрисы изогнуты. Большинство гомозиготных кошек
погибает в раннем возрасте (Шустрова, 1997).
Астения кожи определяется доминантным аллелем Cat.
Проявляется, как недостаток коллагеновых волокон в коже, вследствие чего она
приобретает сильную растяжимость и становится восприимчива к различным
повреждениям (Шустрова, 1997).
.3 Британские короткошерстные и длинношерстные
кошки
.3.1 История британской породы
Британская - одна из самых популярных пород кошек. История
породы берет свое начало с Британских Островов. На них сформировался тип кошки
с круглыми глазами и мордой, с короткой плотной шерстью, хорошо защищающей от
влажного воздуха и насекомых. Бродячий образ жизни сформировал сильное,
крепкое, спокойное духом, миролюбивое животное (Шустрова, 1998).
Селекционная работа над породой началась в конце XIX века.
Англичане решили вывести породу кошек, которая была бы чисто английской: сочетала
бы в себе привлекательную аристократичную внешность и все достоинства
аборигенных короткошерстных кошек. Выведенная порода была названа «Британская
короткошерстная». Порода обязана своим названием Гаррисону Вейеру -
организатору движения любителей кошек (Шустрова, 1998). Впервые британская
порода кошек была представлена на выставке в Лондоне в 1880 году, а первая
родословная была выдана в 1898 году.
После первой мировой войны качественных представителей
британской породы осталось немного. Опытные заводчики пытались восстановить
породу. Большой вклад в сохранение и развитие породы внесла заводчик Кит
Уилсон. Она помогла сохранить породу в годы второй мировой войны (Шустрова,
1998).
Первоначально для этой породы признавался только голубой
окрас (Мей, 2008). В 1950 году американские ассоциации признали британскую
короткошерстную кошку самостоятельной породой и назвали её Британская Голубая.
Другие окрасы продолжали регистрироваться, как Домашние / Американские
Короткошерстные. И только в 1980 году, через 100 лет после первой выставки, CFA
утвердила статус Чемпионства для породы Британская короткошерстная (Шустрова,
1998 г.).
На сегодняшний день британская короткошерстная - это
полностью устоявшаяся порода, которую признают все без исключения
фелинологические организации мира (Шустрова, 1998).
Длинношерстная вариация британской короткошерстной породы
кошек сравнительно недавно была признана в фелинологии, хотя и существует уже
давно. Раньше длинношерстные котята не допускались в разведение. Однако, в
последнее время ситуация изменилась: поклонники длинношерстных британских
кошек, добились признания длинношерстных британцев крупными фелинологическими
ассоциациями, такими как TICA (The International Cat Association) и WCF (World Cat Federation), в других
системах длинношерстные британцы считаются эксперементальной породой
(#"563946.files/image027.gif">
Рис. 27. Черный окрас у британских кошек
Голубой окрас (BRI a). Голубой окрас (Рис.28)
может варьироваться от светло-голубого до средне-голубого. Наибольшим успехом у
экспертов пользуется гамма от средне-голубой до лилово-голубой (Константинова,
2005). Не допускается наличие рисунка. Нос и подушечки лап должны быть
голубыми, а глаза - желтыми, оранжевыми или медными. Шерстный покров у голубых
британцев густой, короткий и мягкий на ощупь («плюшевый») (Константинова,
2005).

Рис. 28. Голубой окрас у британской короткошерстной кошки
Шоколадный окрас (BRI b). Шоколадный окрас (Рис.
29) для британских кошек является сравнительно новым. Он был получен в качестве
побочного продукта при выведении типа колор-пойнт. Селекционеры используют
британских шоколадных для скрещивания с кошками колор-пойнт, так как они
являются носителями гена соответствующего окраса (Константинова, 2005).
Шерстный покров у этих кошек короткий и мягкий,
темно-коричневого цвета. Глаза желтые, оранжевые или медные (Константинова,
2005).

Рис. 29. Шоколадный окрас у британских кошек
Лиловый окрас (BRI c). Лиловый окрас (Рис. 30),
так же как и шоколадный, был получен селекционерами в процессе выведения типа
колор-пойнт. Животные с этим окрасом обладают коротким, густым и мягким
шерстным покровом. Глаза желтые, оранжевые или медные.

Рис. 30. Лиловый окрас у британских кошек
Красный (рыжий) окрас (BRI d)
(Рис. 31) - это равномерный глубокий бриллиантово-красный цвет. Нос и подушечки
лап кирпично-красныме, глаза - золотистые или медныее (Константинова, 2005).

Рис. 31. Красный (рыжий) окрас у британских кошек
Кремовый окрас (BRI e) (Рис.32). Этот
окрас считается одним из наиболее сложных. Чисто-кремовые британцы очень редки.
Шерсть большинства животных с таким окрасом обычно бывает с оттенками и
рисунками, наличие которых, согласно стандарту, исключено (Константинова,
2005). Шерстный покров короткий, густой, мягкий, равномерно окрашенный. Белые
волоски не допускаются. Нос и подушечки лап должны быть розовыми, а глаза -
золотистыми или медными (Константинова, 2005).

Рис. 32. Кремовый окрас у британских кошек
Белый окрас (BRI w) (Рис. 33). Британская
короткошерстная кошка белого окраса бывает трех видов: с сапфирово-голубыми,
золотыми или медными и разноцветными глазами. У животных с разными глазами один
глаз обязательно должен быть сапфирово-голубым, а другой - золотистым, с равной
насыщенностью цвета. Нос и подушечки лап розовые (Константинова, 2005).

Рис. 33. Белый окрас шерсти у британской короткошерстной
кошки
Шерсть должна быть без желтизны. Любые оттенки и пятна
исключаются. Но зачастую белые британцы рождаются с дефектами (отсутствие
слуха, обоняния). Кроме того, не всегда удается получить потомство с безупречно
белой шерстью и голубыми глазами (Константинова, 2005).
Черепаховый окрас. Черепаховый окрас (Рис.
34) может сочетать в себе разнообразные цвета. Основными считаются черепаховый
черный (BRI f), черепаховый голубо-кремовый (BRI g), черепаховый
шоколадно-кремовый (BRI h) и черепаховый лилово-кремовый (BRI j)
окрасы (Константинова, 2005).
Шерстный покров короткий, густой и мягкий. Цвета в окрасе
должны быть равномерно смешаны. Нос и подушечки лап этих кошек розовые или
черные, глаза золотистые или медные (Константинова, 2005).

Рис. 34. Кошка черепахового окраса
Дымчатый окрас. Кошки дымчатых окрасов (Рис. 35), как уже
отмечалось выше, генетически восходят к серебристым. Каждый волосок окрашен в
основной цвет на треть своей длины, нижняя часть, так же как и подшерсток,
чистого серебристого, почти белого цвета. Шерстный покров короткий, густой и
жесткий. Нос и подушечки лап того же цвета, что и шерсть. Глаза дымчатых
британцев должны быть золотистыми или медными (Константинова, 2005).

Рис. 35. Черный дымчатый окрас у британцев
Наиболее распространен черный дымчатый окрас (ns) (Рис.
28). А так же: голубой дымчатый (as), шоколадный дымчатый (bs),
лиловый дымчатый (cs), красный дымчатый (ds), кремовый дымчатый (es),
черный черепаховый дымчатый (fs), голубо-кремовый дымчатый (as),
шоколадно-кремовый дымчатый (hs), лилово-кремовый дымчатый (js)
(Константинова, 2005).
Затушеванные или шиншилловые. Во вторую группу
серебристых окрасов объединяются затушеванные и шиншилловые. Затушеванные и
шиншилловые окрасы выглядят почти белыми, с характерным «напылением» на самых
кончиках шерстинок. У кошек с затушеванным окрасом оно занимает всего лишь
шестую часть волоска, а у особей с шиншилловым окрасом и того меньше - восьмую
(Рис. 36). У кошек с затушеванным окрасом шерсть должна быть затушевана на
спине, боках, голове, ушах и хвосте. Равномерность окраса важнее, чем его
контрастность. Подбородок, живот и нижняя сторона хвоста должны быть как можно
более бледными. Нос и подушечки лап обычно окрашены под цвет шерсти. Глаза у
таких кошек золотистые или медные, без зеленого ободка. Наличие ободка
допускается лишь у серебристых затушеванных (Константинова, 2005).

Рис. 36. Британская короткошерстная затушеванная (Пухова,
2006)
Золотистые (Y). Это большая редкость
для британской породы кошек (Константинова, 2005) (Рис. 37). Подшерсток у таких
кошек не белого, а насыщенного теплого кремового цвета. Шерстинки обычно бывают
с черным или темно-коричневым «напылением» на кончиках. Подбородок, живот и
нижняя сторона хвоста должны быть окрашены в бледно-абрикосовый цвет, нос - в
кирпичный. Подушечки лап черные или темно-коричневые, глаза зеленые
(Константинова, 2005).

Рис. 37. Голубо-золотистая затушеванная кошка
Рисунчатые, или табби (макрель, или полосатый,
пятнистый и мраморный).
Этот окрас является самым разнообразным у британских
короткошерстных кошек (Константинова, 2005). Классический табби характеризуется
плотным четким рисунком с широкими линиями. Окрас лап равномерно-полосатый, с
кольцами, которые плавно переходят в рисунок на теле. Хвост равномерно
окольцован полосками. На шее располагаются несколько замкнутых колец
(«ожерелий»). На лбу присутствует отметина в виде буквы «M». Из уголков глаз
идут непрерывные полоски. Глаза у классического табби должны быть
темно-оранжевого или медного цвета (Константинова, 2005).

Рис. 38. Кошка окраса - тигровый табб
Тигровый табби (Рис. 38) - это плотный четкий рисунок с
узкими линиями. Лапы таких кошек покрыты равномерно распределенными полосами с
тонкими кольцами, которые вверху переходят в рисунок на теле. Хвост полосатый.
«Ожерелья» на шее похожи на цепочки. Так же как у классического табби, на лбу у
тигрового располагается отметина (в виде буквы «M»). От уголков глаз идут
непрерывные полоски. С головы на лопатки опускаются линии, которые на спине
приобретают форму седла. Узкие линии проходят вокруг всего тела. Недостатком окраса
считается наличие на шерсти белых пятен. Глаза медные или темно-оранжевые
(Константинова, 2005).
Для пятнистого табби характерны частые и темные пятна,
которые должны быть хорошо заметны на более светлом фоне. По своей форме они
могут быть округлыми, продолговатыми или похожими на розетку. Голова пятнистого
табби окрашена так же, как у классического табби. Конечности тоже пятнистые. На
хвосте пятен может и не быть, но их наличие желательно. Хвост иногда бывает
украшен разомкнутыми кольцами (Константинова, 2005).
У серебристого табби с рисунком основной цвет шерсти бледный
с явным серебристым оттенком. Рисунок четкий, черный, с отдельными участками,
окрашенными в рыжий цвет. Нос кирпичный, подушечки лап черные и (или)
кирпичные, глаза бриллиантово-зеленые или ореховые (Константинова, 2005).
У коричневого табби основной окрас блестящий,
медно-коричневый. Рисунок черного цвета. Задняя часть лап по всей длине черная,
без рисунка. Нос должен быть кирпичного цвета; подушечки лап - черные или
коричневые, глаза - золотистые или медные (Константинова, 2005).
У голубого табби основной цвет - бледно-голубой или слоновой
кости. Нос и подушечки лап темно-розовые. Глаза золотистые или медные
(Константинова, 2005).
Голубой табби с рисунком (классический, тигровый, пятнистый)
отличается от коричневого наличием на теле и конечностях кремовых пятен или
разводов. Нос и подушечки лап розовые. Глаза золотистые или медные
(Константинова, 2005).
У кремового табби основной цвет - бледно-кремовый. Рисунок
бежевый или кремовый, намного темнее основного цвета, контрастный. Нос и
подушечки лап розовые. Глаза золотистые или медные (Константинова, 2005).
У табби с белым основной цвет - красный, кремовый, голубой
серебристый или коричневый. Согласно требованиям стандарта, чисто белыми у кошки
должны быть морда, «перчатки» на лапках, бедра и нижняя часть туловища. Особое
значение также придается симметричности рисунка (Константинова, 2005) (Рис.
39).
Биколорные окрасы (ван, арлекин и биколор). Биколорные окрасы
представляют собой сочетание белого цвета с каким-либо другим ровным окрасом -
голубым, шоколадным, лиловым, красным, черным, кремовым или любым из
черепаховых. В зависимости от количества белого среди биколорных окрасов
выделяются следующие группы: ван, арлекин, биколор. Кодируются они числами 01,
02 и 03 соответственно (Константинова, 2005).

Рис. 39. Окрас - кремовый мраморный арлекин
Окрас, сочетающий белый и черный цвета, называется «черный
ван», «арлекин» или «биколор», и так далее со всеми остальными цветами.
Стандарт допускает наличие на теле ванов одного или двух окрашенных пятнышек
(Константинова, 2005).
Ситец. Для окраса ситец характерны равномерно
распределенные черные и красные пятна на белом фоне. Стандарт требует, чтобы
трехцветные пятна располагались на макушке, ушах, щеках, хвосте, спине и части
боков животного. Цвета должны быть четкими и блестящими, а пятна - ясно
различимыми. На красных пятнах допускается легкий рисунок табби. Нос и
подушечки лап у кошек этого окраса розовые и (или) черные, глаза золотистые или
медные. Недостатки - несбалансированность цветов и одноцветность лап
(Константинова, 2005).
Разбавленный ситец сочетает в себе равномерно распределенные
кремовые и черные пятна на белом фоне. Трехцветные пятна должны покрывать
макушку головы, уши, щеки, хвост, спину и часть боков кошки. Цвета должны быть
четкими и блестящими, а пятна - ясно видимыми. На кремовых пятнах допускается
легкий рисунок табби. Нос и подушечки лап голубые и (или) розовые. Глаза
золотистые или медные. Недостатками окраса считаются несбалансированность
цветов и одноцветность лап (Константинова, 2005).
Ван ситец - белый окрас с черными и красными (рыжими)
участками на голове, хвосте и лапах кошки. На теле могут быть одно-два
маленьких пятнышка. Глаза должны быть золотистыми или медными (Константинова,
2005) (Рис. 40).

Рис. 40. Окрас ван-ситец
Ван разбавленный ситец - белый окрас с голубыми и кремовыми
участками на голове, хвосте и лапах животного. На теле допускается одно-два
маленьких пятнышка. Глаза золотистые или медные (Константинова, 2005) У кошек
кремово-голубого окраса шерсть светло-голубого тона с кремовыми вкраплениями,
нос и подушечки лап голубые и (или) розовые, глаза золотистые или медные.
Недостатками считаются одноцветность лап, рисунки табби и одноцветные пятна. Голубо-кремовый
окрас может быть только у самок, поэтому потомство с необходимыми
характеристиками можно получить только путем целенаправленного скрещивания
(Константинова, 2005).
2. Материалы и методы исследования
.1 Материалы, использованные в работе
Объекты исследования
В работе использованы кошки британской породы из питомника
«Леонель», (10 особей), а также несколько особей, предоставленных любителями:
кошка Каролина, Ника и коты - Кабачок, Чаки и Чупа (5 особей). Пробы шерсти
кошек были собраны в промежуток времени с января по март 2011 г.
Приборы, инструменты и реактивы
· Бинокулярный микроскоп «Бимам»
· Микроскоп «Микмед»
· Гребень для вычесывания шерсти
· Предметные стекла
· Канадский бальзам
· Скотч
· Фотокамера Nikon E4500
· Пинцет
· Скальпель.
2.2 Методы, использованные в работе
Пробы шерсти забирали с боков и со спины кошки. Небольшой
пучок вычесывался с помощью тонкого гребня. Собранные образцы волос
каталогизировали.
Первоначальный осмотр волоса производился под лупой (х 7).
Оценивалась толщина, длина волоса, а так же текстура (прямой или завитой) и
цвет волоса.
Для микроскопического исследования волосы фиксировались в
канадском бальзаме и помещались на предметные стекла (Голубева, 2008 г.).
Длинные волосы разрезались на части. Анализ препаратов производился на
микроскопе «Микмед» (увеличение 8х). Затем волосы снимали на камеру.
Микроскопически оценивалась струкрура и распределение пигмента в корковом слое
волоса, центральном (сердцевинном), на кончике волоса и в основании.
Оценивались как остевые, так и пуховые волосы. Определялся фенотип, генотип и
распределение пигмента по волосу. Результаты заносились в сводную таблицу.
Образцы волос оценивались по следующим критериям:
· Толщина волоса (толстый или тонкий волос)
· Длина волоса (короткий, средней длины,
длинный)
· Распределение пигмента по длине волоса
(типпинг) ((а- неокрашенный, b - типированный (шиншилла), с -затушеванный, d-
дымчатый, е - окрашенный полностью). (Рис. 42)
· Цвет (а) пигмента
· Когда вырвали волос?
· Текстура волоса (завитая, волнистая или
прямая)
· Равномерное распределение пигмента,
разряженное распределение пигмента (Рис. 41) (Голубева, 2008г.; Мычко, Сотская,
Маклакова).
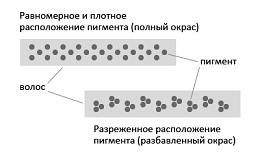
Рис. 41. Распределение пигмента по волосу кошки
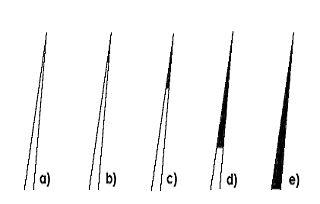
Рис. 42. Распределение пигмента по длине волоса или типпинг
(а- неокрашенный, b - типированный (шиншилла), с -затушеванный, d- дымчатый, е
- окрашенный полностью)
3. Результаты и обсуждение
.1 Особенности структуры и пигментации волос у
кошек британской породы
В ходе выполнения работы была собрана и каталогизирована
коллекция волос кошек британской породы. Всего в период с января 2010 по март
2011 года удалось собрать образцы волос, вычесанные со спины и с боков у 17-ти
кошек британской породы.
Большинство исследованных кошек (13 особей) были
короткошерстными, часть (4 особи) - длинношерстными.
Преимущественно в выборку вошли кошки голубого окраса (12
особей), который считается «классическим» у британской породы, но также были
исследованы и кошки лилового (4 особи) и рыжего (1 особь) окраса.
По возможности для исследования были использованы образцы как
остевых, так и пуховых волос.
Визуальный (с использованием бинокулярной лупы) анализ
позволил оценить длину, текстуру, цвет и примерное распределение пигмента по
длине волоса.
Распределение пигмента по отдельным частям волоса
(центральной, на кончике и у основания), характер упаковки пигмента,
особенности коркового слоя, оценивались при микроскопическом исследовании.
Полученные данные (см. приложение) позволили определить
наиболее вероятные алели генов окрасок, носителями которых являются
исследованные особи. При этом учитывались только гены, имеющие наибольшее
селекционное значение для породистых кошек - а (ген, обуславливающий
равномерное или зонарное распределение пигмента по частям волоса), B (ген, определяющий цвет
основного пигмента), dl (ген, определяющий наличие или отсутствие
пигмента на кончике волоса), I (ген, обуславливающий неокрашенность основания
волоса у длинношерстных особей), d (ген, обуславливающий ослабление окраски
вследствие разреженного расположения пигмента).
В результате были составлены генотипы исследованных
британских кошек по генам окрасок. Результаты исследования представлены в
сводной таблице 11.
Таблица. 11. Сводная таблица результатов исследования волос
кошек британской породы
|
Фенотип
волоса
|
Распределение
пигмента по волосу
|
|
Окрас
|
Остевой
волос
|
Пуховой
волос
|
По длине
волоса
|
В центре
волоса
|
На кончике
волоса
|
Корковый
слой
|
|
Длина
|
Структура
|
Длина
|
Структура
|
|
|
|
|
|
Голубой-дымчатый
|
Длинный
|
Волнистый,
тонкий
|
Короткий
|
Волнистый,
тонкий
|
Не равномерное
|
Разреженное
|
Не равномерное,
есть просветы без пигмента
|
Прозрачный
|
|
Голубой
|
Короткий
|
Прямой, средней
толщины (ближе к толстому).
|
Короткий
|
Волнистый,
тонкий
|
Не равномерное;
У основания и на кончике отсутствует
|
Разреженное
|
Не равномерное,
на кончике отсутствует
|
Прозрачный
|
|
Голубой
|
Средней длинны
(ближе к короткому)
|
Прямой, тонкий
|
Короткий
|
Волнистый,
тонкий
|
Не равномерное;
У основания менее выражен, на кончике отсутствует
|
Равномерное в
центре остевого волоса, разреженное в центре пухового.
|
Не равномерное,
на кончике отсутствует
|
Прозрачный
|
|
Голубой
|
Средней длинны
(ближе к короткому)
|
Прямой, тонкий
|
Короткий
|
Волнистый,
тонкий
|
Не равномерное;
На кончике пигмента нет
|
Разреженное
|
Не равномерное,
пигмент выражен слабее или вообще отсутствует
|
Прозрачный
|
|
Голубой
|
Длинный
|
Волнистый,
тонкий
|
Длинный
|
Волнистый,
тонкий
|
Не равномерное;
На кончике пигмента нет
|
Равномерное в
остевом волосе, не равномерное в пуховом (есть участки без пигмента)
|
Не равномерное,
на кончике отсутствует
|
Прозрачный
|
|
Голубой
|
Длинный
|
Прямой, средней
толщины (ближе к толстому)
|
Длинный
|
Волнистый,
тонкий
|
Не равномерное;
На кончике пигмента нет
|
Равномерное в
остевом волосе, разреженное в пуховом.
|
Не равномерное,
на кончике отсутствует
|
Прозрачный
|
|
Голубой
|
Короткий
|
Прямой. Тонкий
|
Короткий
|
Волнистый,
тонкий
|
Не равномерное;
На кончике пигмента нет
|
Равномерное в
остевом волосе, разреженное в пуховом.
|
Не равномерное,
на кончике отсутствует
|
Прозрачный
|
|
Голубой-дымчатый
|
Короткий
|
Прямой, толстый
|
Короткий
|
Волнистый,
тонкий
|
У пухового
волоса - не равномерное, на кончике пигмента нет. У остевого - относительно
равномерное, на кончике пигмент выражен слабее.
|
Равномерное в
остевом волосе, разреженное в пуховом.
|
У пухового
волоса полное отсутствие, у остевого волоса есть, но выражен слабее
|
Прозрачный
|
|
Голубой-дымчатый
|
Длинный
|
Волнистый,
тонкий
|
Длинный
|
Волнистый,
тонкий
|
Не равномерное;
У основания и на кончике отсутствует.
|
Разреженное
|
Не равномерное.
Пигмента меньше, чем в других частях волоса
|
Прозрачный
|
|
Голубой-дымчатый
|
Длинный
|
Прямой, толстый
|
Короткий
|
Волнистый,
тонкий
|
У пухового
волоса - не равномерное, на кончике пигмента нет. У остевого - относительно
равномерное, на кончике пигмент выражен слабее.
|
Равномерное в
остевом волосе, разреженное в пуховом.
|
Не равномерное.
У пухового волоса полное отсутствие, у остевого волоса есть, но выражен
слабее
|
Прозрачный
|
|
Голубой
|
Короткий
|
Прямой, тонкий
|
Короткий
|
Волнистый,
тонкий
|
Не равномерное;
На кончике пигмента нет
|
Разреженное
|
Не равномерное,
отсутствует
|
Прозрачный
|
|
Голубой
|
Короткий
|
Прямой, тонкий
|
Короткий
|
Волнистый,
тонкий
|
Не равномерное;
На кончике пигмента нет
|
Разреженное
|
Не равномерное,
отсутствует
|
Прозрачный
|
|
Лиловый
|
Короткий
|
Прямой, средней
толщины (ближе к тонкому)
|
Короткий
|
Волнистый,
тонкий
|
Не равномерное;
На кончике пигмента нет
|
Равномерное в
остевом волосе, разреженное в пуховом.
|
Не равномерное,
на кончике отсутствует
|
Прозрачный
|
|
Лиловый
|
Короткий
|
Прямой, средней
толщины (ближе к тонкому)
|
Короткий
|
Волнистый,
тонкий
|
Не равномерное;
На кончике пигмента нет
|
Равномерное в
остевом волосе, разреженное в пуховом.
|
Не равномерное,
на кончике отсутствует
|
Прозрачный
|
|
Лиловый
|
Короткий
|
Прямой, тонкий
|
Короткий
|
Волнистый,
тонкий
|
Не равномерное;
На некоторых участках и на кончике отсутствует.
|
Равномерное в
остевом волосе, разреженное в пуховом.
|
Не равномерное,
на кончике пухового волоса отсутствует, на кончике остевого волоса выражен
слабее
|
Прозрачный
|
|
Лиловый
|
Короткий
|
Прямой, тонкий
|
Короткий
|
Волнистый,
тонкий
|
Не равномерное
|
Разреженное
|
Не равномерное,
отсутствует
|
Прозрачный
|
|
Рыжий
|
Коротий
|
Прямой, тонкий
|
Короткий
|
Волнистый,
тонкий
|
Не равномерное;
На кончике пигмента нет
|
Равномерное в
остевом волосе, разреженное в пуховом
|
Не равномерное,
отсутствует
|
Прозрачный
|
Из таблицы 11 видно, что из 17 исследованных кошек 12 имели
голубую окраску 9 (из них 4 голубую-дымчатую), 4 лиловую и 1 рыжую. Таким
образом, наибольшая часть выборки представлена кошками голубого окраса. Это
соотношение отражает и популярность кошек различных окрасов у любителей (Якобс,
2008). Так в большинстве питомников 70% предложений составляют котята именно
голубого и голубого дымчатого окрасов (Шевченко, 2006).
По длине шерсти - большинство - короткошерстные. По структуре
волос - остевой волос чаще прямой, а пуховой - волнистый. Хотя у некоторых из
исследованных длинношерстных кошек образцов (2, 3, 7) и остевой волос
волнистый. У всех исследованных кошек волосы тонкие.
Голубая - дымчатая окраска наблюдалась только у
длинношерстных кошек, под воздействием гена I, который обесцвечивает
основание волоса. У большинства короткошерстных голубых и лиловых кошек, кончик
волоса лишен пигмента. Это происходит под воздействием гена dl.
Обнаружились разнообразные варианты распределения пигмента по
волосу. В центре пухового волоса всегда - разреженное, а в центре остевого
преимущественно - равномерное (9 образцов), но есть и разреженное (7 образцов).
На кончике и у основания волоса не равномерное распределение пигмента
наблюдается у всех кошек. На кончике остевых волос 3 кошек (11, 13, 14) пигмент
есть, но выражен слабее, чем в центре волоса. У остальных 15 образцов на
кончиках остевых волос пигмента нет. У всех образцов на кончиках пуховых волос
нет пигмента. Корковый слой у всех прозрачный.
Таким образом, в большинстве случаев в генотипе исследованных
британских кошек присутствуют гены:
d, обуславливающий разреженное распределение
пигмента в различных частях волоса. При этом зачастую ген проявляется только в
остевом и пуховом волосах (образцы 1, 2, 3, 4, 15, 17) или только в пуховом
(образцы 5, 8, 9, 10, 11, 12, 13, 14, 16) волосе.
В результате фенотипический эффект этого гена также различен
и проявляется в разной насыщенности (интенсивности) окраски.
a, обуславливающий равномерное распределение
пигмента по волосу (1-17 образцы). Его доминантный аллель А обычно вызывает
зонарное распределение пигмента по волосу с чередованием не менее 3-х полос,
либо одновременное обесцвечивание кончика одних волос и основания других. Таких
вариантов у исследованных кошек не выявлено.
У большинства же исследованных животных наблюдалось
ослабление окраски либо в кончике, либо в основании волоса. Такой
фенотипический эффект может быть вызван присутствием в генотипе генов dl, I.
- dl, обуславливающий обесцвечивание кончика
волоса.
У исследованных кошек, имеющих неокрашенный или слабо окрашенный
кончик волоса наблюдается постепенное ослабление окраски на кончике. То есть
присутствует менее окрашенная «переходная зона» между окрашенной частью волоса
и неокрашенным кончиком (см. приложение).
При этом неокрашенный кончик волоса присутствует в основном у
короткошерстных кошек.
I, обуславливающий обесцвечивание основания
волоса. Выявлен в основном у длинношерстных кошек. Соответственно, рецессивный
аллель этого гена - i - у короткошерстных. Однако, такое наблюдение может
свидетельствовать и о том, что ген I имеет разную экспрессивность в волосе разной
длины и не всегда проявляется в коротком волосе. Однако это предположение на
фенотипическом уровне нам не удалось проверить.
B и b. Кошки голубого и дымчатого окрасов
содержат в генотипе аллель В, обуславливающую синтез в организме
эумеланина, а лиловые и рыжий кот - аллель b, характеризующуюся
наличием в организме феумеланина (образцы 1, 9, 10, 11, 16). При этом
ослабление окраски и формирование красивых голубых, лиловых, дымчатых и рыжих
окрасов, связана я деятельностью гена d.
Анализ результатов, приведенных в таблице 11, показывает, что
даже кошки одного типа окраски составляют генотипически разнородную группу. Так
у кошек голубой окраски, но разной длины волоса обнаружились разные
распределения пигмента.
У длинношерстных в основном обесцвечено основание волоса, а у
короткошерстных - кончик.
Для более точного анализа генотипов окрасок, характерных для
кошек британской породы, необходим анализ большей выборки из большего числа
питомников, в которых кошки имеют различное происхождение.
Сложность сбора большей выборки обусловлена тем, что
питомники неохотно разрешают исследовать своих кошек, особенно если речь идет
об образцах шерсти. Также практически невозможно получить достоверную
информацию о генотипах окрасов родителей. Поэтому, остается невыясненным,
являются ли исследованные породистые кошки гомозиготными по некоторым из генов
окрасок.
Окраска для большинства домашних животных является важнейшим
признаком породы (Сотская, 2010). И селекция по этому признаку у собак,
например, ведется не менее тщательная, чем по другим экстерьерным признакам.
У кошек, этот признак часто рассматривается заводчиками как
второстепенный. Особенно это касается молодых пород, к которым относится и
британская. Заводчики часто не хотят обращать внимание на передачу генов
окрасов в родословных разводимых кошек. Часто это связано с элементарным
незнанием генетики окрасок млекопитающих и механизмов, влияющих на
пигментогенез (Шустова, 1997). Таким образом, даже кошки из питомников не
всегда имеют точно определенный генотип относительно генов окрасок. Возможно, с
этим и связано неохотное представление заводчиками информации по животным их
питомников.
Безусловно, такая ситуация требует разработки системы
тестирования кошек по генам окрасок. Только кошка с четко определенным
генотипом по экстерьерным, важным для породы признакам, может считаться
породистой. Тестирование по генам окрасок может проводиться как на
фенотипическом уровне (путем визуальной оценки, микроскопического исследования
волос, составления родословных с учетом известных для млекопитающих генов
окрасок), так и на уровне генотипа. Последнее, конечно, требует разработки
специальных тест-систем, анализирующих конкретные гены окрасок.
Выводы
. У кошек британской породы дымчатый окрас формируется под
влиянием генов dl и I, обуславливающих ослабление окраски или неокрашенность либо
основания, либо кончика волоса. Действие этих генов проявляется как в пуховом,
так и в остевом волосах.
. У исследованных кошек британской породы классические окрасы
(голубой, лиловый) обусловлены присутствием в генотипе гена d, определяющего
разреженное расположение пигмента в волосе.
. Британские кошки одной фенотитипической группы (голубые,
лиловые, дымчатые) по генам окрасов являются генотипически гетерогенной
группой: в их генотипах присутствуют различные аллели одних и тех же генов
окрасов. Это указывает на пренебрежение заводчиками признаком окраски как
селекционно-важным для породы.
Список литературы
1. Алиев
Г.А., Рачковский М.Л. 1986. Взаимодействие регуляторных генов А и Е и
генов-модификаторов в процессе ингибирования меланогенеза у овец шерстных пород
// Доклады АН СССР. М.,, т. 287, С. 990-994.
2. Алиев
Г.А., Рачковский М. Л. 1986b. Генетика пигментации шерстного покрова овец //
Животноводство. №8, C. 32-34.
. Алиев
Г.А, Рачковский М.Л. 1989. Исследование процесса включения меланоцитов в волосы
ягнят разных окрасок, Доклады ВАСХНИЛ, № 9, С. 178-185.
. Бородин
П.М. 2009, Кошки и гены, М.: Книжный дом «ЛИБРОКОМ», С. 132
. Бородин
П.М., 2009, Кошки и гены: 30 лет спустя //, №4, С. 89-97
. Ватти
К.В., Алексеевич Л.А. 1976. Сравнительная генетика и онтогенетика окрасок у
животных //Физиологическая генетика, Ленинград, С. 326-350.
7. Голубева
О.Н. Британская порода кошек // Питомник британских кошек «SOFTCAT»,
<http://www.softcat.ru/britishcat.html>.
. Голубева
Н.А. 2008. Частоты генов окраса, длины шерсти, и спектр врожденных аномалий в
современных популяциях кошек. Диссертация канд. биол. наук. С. 185
. Гилл Д.
2001, Кошки: Мини энциклопедия (все и обо всем). М: «Астрель», С. 260
. Деревянко
В. 2008. Планета кошек: Британские короткошерстные (фильм), ПТФ Сейприс.
. Е.
Друбецкая, 2010. Кто кому хозяин, или Кошка в Древнем Египте // Мой друг кошка,
№12, С. 16-17
. Константинова
Е.А. 2005. Домашние кошки, М.: «Вече», С. 325
. Коростовцев
М.А., 2000. Религия древнего Египта, СПб.: Летний сад, С. 334
. Красичкова
А.Г. 2006. Длинношерстные кошки, М.: «Вече», С. 160
. Мей Д.
2008. Все о породах кошек, СПб.: Кристалл, С. 176
. Мычко
Е.Н., Сотская М., Маклакова Л. К. 2005. Генетика окрасов в породе
среднеазиатская овчарка: микроскопический анализ пигментации остевого волос //
Научный сборник РФСС, С. 5-6
. Нестерова
Д.В., 2005. Кошки-сфинксы. М.: «Вече», С. 64
. Пинтер
А. 1993. Кошки, коты, котята. М.: ЭКОЦЕНТР - ВНИРО, С. 272
. Поллард
М.. 2009. Энциклопедия. Кошки. М.: АСТ, Астрель, Харвест, С. 384
. Прасолова
A., Трут H, Всеволодов З.Б., Латыпов И.Ф. 1989. Феногенетический анализ
некоторых изменений окраски меха у серебристо-черных лисиц, возникших в
процессе доместикации // Генетика. т. 25, С. 1626-1634.
. Робинсон
Р. 1995. Генетика окрасов собак. М.: OCR, - 88
с.
. Сингер
М., Берг П. 1998. Гены и геномы, М.: Мир. В 2-х т.
. Сотская
М.Н., Московкина Н. Н.. 2000. Генетика и наследственные болезни собак и кошек.
М.: Аквариум, С.448
. Сотская
М.Н. 2010. Генетика окрасов и шерстного покрова собак, М.: Аквариум-Принт, С.
328
. Фирсова
Е.В. 2006. Кошки от А до Я, М.: Вече, С. 320
. Шварцман
П.Я. 1986. Полевая практика по генетике с основами селекции. М.: Просвещение. -
С. 111
. Шевченко
Е.А. 2006. Все о британских кошках, М.: «Аквариум», С. 224
. Шустрова
И. 1997. Кошки: Генетика и племенное разведение, М.: Мир, - С. 128
. Шустрова
И.В. 2002. От генетики к разведению // «Друг», № 10-11, С. 8-10
. Якобс Л.
Британские короткошерстные, http://ridolina.mybb.ru/ viewtopic.php? id=31
<http://ridolina.mybb.ru/ viewtopic.php?id=31>
. Панченко
М. 2008. Британская длинношерстная, <http://www.mysticat.ru/stati22.htm>.
. http://innos01.deviantart.com/journal/17953348/#/d1blsp4
- кошка черного окраса
. http://prodam.slando.ru/moskva/klubnaya_britanskaya_devochka_redchayshego_okrasa_tsinnamon_P_22490089.html
- кошка окраса циамон
. http://www.zoopets.ru/view/?
6982 <http://www.zoopets.ru/view/?6982> - кошка лилового окраса
. http://forum.plasticsur.ru/index.php?
s=d0edc6f33357bcd12fafc3999356455a&showtopic=5098&st=500 - кошка
черного - дымчатого окраса
. <http://www.baby.ru/community/view/920867/forum/post/9820608/>
- кошка белого окраса
. ---http://egida.by/publ/90101830
<http://egida.by/publ/90-1-0-1830> - кошка рыжего окраса
. http://rasty.kiev.ua/viewtopic.php?
t=13862&start=0
<http://rasty.kiev.ua/viewtopic.php?t=13862&start=0> - кошка
абессинского окраса
. <http://www.nodemetra.lv/Icula.html>
- кошка окраса камео
. <http://razdelanimals.biz-market.ru/number0924730.html>
- кошка окраса - серебристая шиншилла
. http://marfoto.ucoz.ru/photo/47-0-682
- кошка окраса - золотистая шиншилла
. <http://www.dmir.ru/mo/pets/cats/225420/>
- кошка окраса - полосатый тебби
. <http://www.magicbox-cats.ru/pomet-archive.html>
= кошка окраса - пятнистый тебби
. http://pesikot.org/forum.archive/index.php?
showtopic=95759 <http://pesikot.org/forum.archive/index.php?showtopic=95759>
- кошка черепахового окраса