ْÑ,
êðèñòàëëè÷íîñòü
50-80%,
ìîëåêóëÿðíàÿ
ìàññà 100-800 кДа.
Преимущества разработки ПГА заключается в том, что они нетоксичны и
биосовместимы; разрушаются в биологических средах до конечных продуктов (СО2
и Н2О); обладают антиоксидантными свойствами и пьезоэлектрическим
эффектом; термопластичны, перерабатываются в изделия (пленки, полые формы,
нити) из порошков, растворов и расплавов; не требуют технологических добавок;
подлежат стерилизации общепринятыми методами; для синтеза используют доступные
и дешевые отечественные реагенты.
Основное ограничение для коммерческой
эксплуатации бактериального ПГА - высокая промышленная стоимость относительно
этого для получения нефтяных товарных пластмасс, типа полиэтилена. Именно в
этой перспективе синтез ПГА в генетически проектируемых предприятиях был
замечен как перспективный подход для производства разлагаемых микроорганизмами
полимеров на крупном масштабе и за низкую цену [15].
Среди охарактеризованных к настоящему времени
полигидроксиалканоатов выделено несколько групп полимеров. Основные структуры
ПГА можно иллюстрировать следующей схемой, показанной на рис.1:
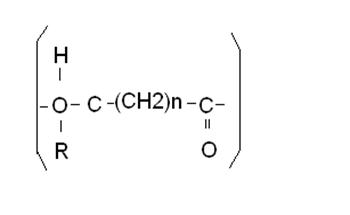
n=1 R=
водород поли (3-гидроксипропионат)
R= метил поли
(3-гидроксибутират)
R= этил поли
(3-гидроксивалерат)
R= пропил поли
(3-гидроксигексаноат)
R= пентил поли
(3-гидроксиоктаноат)
R= нонил поли
(3-гидроксидодеканоат)
n=2 R=
водород поли (4-гидроксибутират)
n=3 R=
водород поли (5-гидроксивалерат)
Рис.1. Основные типы структуры
полигидроксиалканоатов
1.2 Метаболические пути
синтеза
На примере наиболее изученного из ПГА - полимера
β-оксимасляной
кислоты установлено, что пути его синтеза практически одинаковы у различных
микроорганизмов (Alkaligenes,
Azotobakter, Pseudomonas).
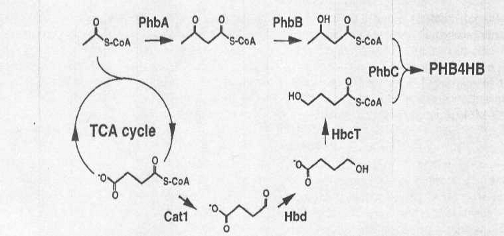
Рис.2. Биосинтез P (3HB-4HB)
в рекомбинантном E. coli
при использовании гетерологичных генов от Clostridlum
kluyveri.
HB мономер в синтезе P (3HB-4I
IB) получен из сукцината. В сукцинате преобразовывают 4HB-CoA
ферменты, которые образовывают дугу закодированный генами от
грамположительного, строго анаэробного C. kluyveri
микроба.
В биосинтезе
полигидроксиалканоатов ключевыми являются три фермента: β-кетотиолаза,
ацетоацетил-КоА-редуктаза и ПГА-синтаза, которые кодируются соответственно
генами phbA, phbB,
phbC (Рис.3). Регуляция
процесса синтеза ПГА может осуществляться на нескольких уровнях: на уровне
экспрессии генов специфическими факторами среды (например, недостаток
питательных элементов) или на уровне регуляции активности ферментов
специфическими клеточными компонентами, являющимися их субстратами или
ингибиторами. Возможен смешанный тип регуляции активности ферментов,
участвующих в метаболизме ПГА. В самых первых работах, посвященных изучению
механизма синтеза ПГА, было показано, что регуляция синтеза ПГА у R.eutropha
осуществляется метаболитами на ферментативном уровне при ведущей роли
внутриклеточной концентрации свободного коэнзима А, который ингибирует
кетотиолазную реакцию у A.eutrophus,
у Azotobacter
beijerinckii, а также у Zoogloea
ramigera. Установлено, что
синтез ПГА стимулируется высокой внутриклеточной концентрацией НАДФН и высоким
соотношением НАДФН/НАДФ.
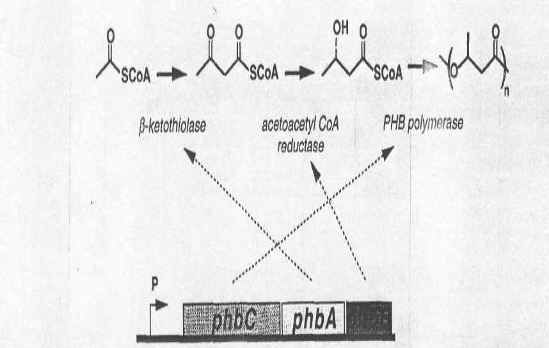
Рис.3. Схематичное изображение
генов ферментов синтеза ПГА и продуктов их экспрессии.
β-кетотиолаза
является первым ферментом синтеза полимера и катализирует конденсацию двух
молекул ацетил-КоА, фермент также катализирует последнюю реакцию в процессе
эндогенной деградации полимера. Таким образом, этот фермент играет ключевую
роль в циклическом метаболизме полимера. Активность фермента не ингибируется
физиологическими концентрациями метаболитов клетки, но реакция конденсации
резко снижается при накоплении свободного КоА. Эффект ингибирования свободным
КоА синтеза полимера является очень важным аспектом метаболической регуляции
клетки, так как в условиях повышенных концентраций свободного КоА субстраты
используются главным образом в биосинтетических реакциях, приводящих к синтезу
белков, нуклеиновых кислот и других первичных макромолекул. При этом ацетил КоА
используется в основном цикле трикарбоновых кислот (ЦТК), где на первом этапе
цикла цитратсиназа высвобождает свободный КоА. В этих условиях внутриклеточная
концентрация свободного кофермента значительно выше, чем в условиях
несбалансированного роста. В результате высокий уровень КоА ингибирует
активность β-кетотиолазы и в
конечном итоге,- синтез полимера. О регуляторной роли ЦТК в направлении потоков
ацетил КоА в клетке свидетельствуют данные, полученные на мутанте R.eutropha,
лишенного изоцитратдегидрогеназной активности, который синтезирует со
значительно большей скоростью, чем дикий штамм. Вместе с тем, показано, что
активность первых двух ферментов ЦТК ингибируется повышенными концентрациями
восстановленных никотинамидных нуклеотидов при несбалансированном росте. Таким
образом, синтез полимера, например, в R.eutropha
начинается, когда ингибируются активности ферментов ЦТК (цитратсиназы и
изоцитратдегидрогеназы). Это приводит к увеличению внутриклеточной концентрации
ацетил-КоА, восстановленных пиридиннуклеотидов, снижению концентрации
свободного кофермента А и окисленных форм нуклеотидов. Сходный контроль
ферментативной активности на уровне метаболитов был подтверждён на
рекомбинантном штамме E.coli,
трансформированном генами синтеза полимера из R.eutropha.
На первых порах в R.eutropha
было обнаружено две β-кетотиолазы.
Уровень ферментативной активности кетотиолаз в клетках, выращенных в условиях
лимитирования роста азотом или углеродом, одинаков (0,20 и 0,27 U/мг
белка, соответственно). Это позволило авторам сделать вывод о конститутивной
природе фермента. Было установлено, что две кетотиолазы отличаются субстратной
специфичностью. Так β-кетотиолаза
А активна с ацетоацетил-КоА и 3-кетопентаноил-КоА и является главным
претендентом на участие в синтезе субстратов для ПГА. Субстратная специфичность
β-кетотиолазы
В оказалась более широкой, фермент активен с длинноцепочечными 3-кетоацильными
производными КоА от 4 до 10 атомов углерода. Было высказано предположение, что
эта кетотиолаза является ферментом β-окисления
жирных кислот. Участие кетотиолазы В в синтезе субстратов для образования
полимера было подтверждено на рекомбинантных штаммах Escherichia
coli, дефектным по
генам, ответственным за деградацию жирных кислот и транспорт пропионата, и
содержащим оперон синтеза полимера из A.eutrophus
без гена кетотиолазы (phaA).
В дальнейшем из R.eutropha
были клонированы три последовательности, кодирующие три β-кетотиолазы.
Было установлено, что β-кетотиолаза,
кодируемая phaA геном,
является β-кетотиолазой А и
высокоактивна с ацетоацетил-КоА. Вторая β-кетотиолаза,
являющаяся продуктом bktB
гена, отвечает преимущественно за синтез кетовалерил-КоА и в меньшей степени -
кетогексаноил-КоА, а также, возможно, соответствует β-кетотиолазе
В и участвует в β-окислении
коротко- и среднецепочечных жирных кислот. Функция третьей кетотиолазы пока до
конца не выяснена, но она также активна с ацетоацетил-КоА как субстратом.
Участие β-кетотиолазы
В в синтезе полимера было подтверждено в работе. Авторы установила, что в
условиях блокирования β-окисления
жирных кислот в клетках R.eutropha
создается пул среднецепочечных 3-гидроксикислот, служащих субстратом для
синтеза ПГА. В результате клетки накапливают гетерополимерные ПГА с включением
гидроксигексаноата до 7-10 мол.%.
Известно, что пул ацетил-КоА
(основного субстрата β-кетотиолазы)
формируется в клетках R.eutropha
в результате катаболизма жирных кислот, аминокислот и углеводов. В условиях дефицита
азота в среде в клетках синтезируется гомополимер оксимасляной кислоты. Синтез
гетерополимерных ПГА обычно наблюдается при добавлении в среду жирных кислот
или других альтернативных ко-субстратов. Однако, мутант A.eutrophus
с измененным анаболизмом разветвленных аминокислот синтезировал
многокомпонентные ПГА без добавок в культуральную среду жирных кислот. Это
объясняется тем, что при катаболизме валина, треонина и изолейцина образуется
пропионил-КоА, вступающий в реакцию конденсации с ацетил-КоА, при этом в
кетотиолазной реакции образуется 3-кетовалерат-КоА, восстанавливающийся далее
до соответствующей гидроксикислоты - субстрата ПГА-синтазы.
Таким образом, в R.eutropha
обнаружено, по крайней мере, три кетотиолазы, отличающейся своей специфичностью
к субстратам. Продукты всех кетотиолаз используются в синтезе ПГА. Фермент
играет ведущую роль в клеточном метаболизме, так как именно он, реагируя на
уровень ацетил-КоА и КоА в клетке, включает синтез запасного материала - ПГА.
На втором этапе синтеза ПГА
происходит восстановление продуктов, полученных в кетотиолазной реакции, до
гидроксикислот. Это реакция катализируется вторым ферментом биосинтетического
пути ПГА - ацетоацетил-КоА-редуктазой. В R.eutropha
обнаружено две редуктазы - НАДН - и НАДФН-зависимые. В синтезе полимера
участвует только НАДФН-зависимая ацетоацетил-КоА редуктаза. Фермент отличается
высокой специфичностью к длине цепи кетоацил производных КоА и активен с
3-гидроксиацил-КоА с длиной цепи от 4 до 6 атомов углерода. Высокая специфичность
редуктазы из R.eutropha
к длине цепи субстрата была подтверждена в более поздних работах с
рекомбинантными штаммами Е. coli.
Критическим фактором, определяющим активность фермента, является соотношение
НАДФН/НАДФ. При высоких значениях этого соотношения активность фермента
возрастает. Установлено, что скорость синтеза полимера в R.eutropha,
выращиваемого на различных источниках углерода, строго зависит от концентрации
в клетке НАДФН. Эти данные подтверждают ранее выдвинутое предположение о том,
что редуктаза определяет скорость синтеза полимера в клетке.
Третьим ключевым ферментом
биосинтеза, определяющий тип ПГА, является ПГА - синтаза.
Таким образом, в синтезе
полигидроксиалканоатов участвуют три фермента. Первые два отвечают за синтез
субстратов, причем кетотиолаза включает синтез полимера. ПГА - синтаза
определяет количество и качество синтезированного ПГА.
1.3
Свойство и структура ПГА - синтазы, выделенной из R.eutropha
В настоящее время выделено и
охарактеризовано более 50 структурных генов синтаз из разных микроорганизмов.
ПГА - синтаза из R.eutropha
состоит из одного типа субъедениц с молекулярной массой около 64 кДа и
кодируется геном phbC.
В клетке ПГА - синтаза находится в двух формах: растворимой и связанной с
гранулами полимера. Обе формы наиболее активно используются для построения
полимерных молекул короткоцепочечных (от 3 до 5 атомов углерода)
3-гидроксиацил-КоА и это свидетельствует о строгой специфичности фермента к
короткоцепочечным производным. Эти данные были подтверждены в системе синтеза
полимера in
vitro. ПГА - синтаза,
выделенная из рекомбинантного штамма Е. coli,
с трансформированным ПГА - синтазным геном из R.eutropha,
высокоактивна с 3-ОН-С4-КоА и 3-ОН-С5-КоА субстратами. С
3-ОН-С6-КоА активность фермента резко снижается и составляет всего
0.94% от таковой с гидроксибутират-КоА. Наибольшее сродство фермент имеет по
отношению к кислотам с ОН-группой в β-положении.
Перемещение ОН-группы к метильному концу снижает способность субстрата
связываться с ферментом, однако не влияет на скорость синтеза полимера. Также
доказано, что локализация гидроксила у карбоксильного конца полностью
дезактивирует фермент. К настоящему моменту показано, что субстратная
специфичность ПГА - синтазы in
vivo может существенно
отличаться от ее характеристик in
vitro. ПГА - синтаза in
vivo может включать в
полимер в небольших количествах и более длинные гидроксикислоты - С6,
С8 и С10. Это свидетельствует о более широком спектре
субстратов, используемых синтазой для построения полимеров. Разница в
субстратной специфичности фермента in
vivo и in
vitro может быть связана
со сроками наблюдения за включением субстратов в ПГА, или разницей в
специфической среде окружения фермента. Полученные данные свидетельствуют о
конститутивном синтезе ПГА-синтазы. Например, в условиях лимита по углероду R.eutropha
синтезирует преимущественно растворимую форму фермента, а при переносе культуры
в условия дефицита азота, активность растворимой ПГА-синтазы резко падает, и в
клетках в основном определяют гранулозависимую форму фермента. При этом
суммарная активность фермента остается на одном и том же уровне (0.05 u/mg
protein). Растворимую
форму ПГА-синтазы в достаточном количестве выделили из рекомбинантного штамма
Е. coli, который содержал
ПГА - синтазный ген. Рекомбинант оказался способным синтезировать фермент с
высоким выходом, но так как в среде отсутствовали субстраты, в клетках
накапливалась только растворимая форма фермента. Молекулярный вес растворимой
синтазы составляет 64 кДа. Кинетический анализ показал, что реакция полимеризации
3-ОН-С4-КоА с этим ферментом начинается после достаточно
продолжительной лаг фазы, которая увеличивалась при снижении концентрации
фермента в реакционной смеси. Эти наблюдения позволили предположить, что для
сокращения длительности лаг фазы необходим праймер, который позволил бы
переводить фермент из мономерного в димерное состояние. И, действительно,
использование в реакции полимеризации олигомера 3-ОН-С4 исключило
лаг фазу, а в реакционной смеси при этом обнаружили мономерные и димерные формы
фермента. К настоящему времени доказано, что только димерная форма фермента
обладает каталитической активностью. Это хорошо согласуется с механизмом
реакции полимеризации, согласно которому в реакцию полимеризации должны
включаться две тиольные группы. Однако у ПГА - синтазы обнаружен только один
цистеин в положении 319, отвечающий за образование полимерных цепей. Достаточно
много было выдвинуто предположений о посттрансляционных модификациях фермента
пантотеновой кислотой, которая могла бы ввести вторую тиольную группу для
успешной работы фермента. Однако наиболее доказательной оказалась модель
образования активного фермента в виде димера, у которого две тиольные группы
расположены в Cys319,
локализующие в консервативном участке субъединиц. На основании этих данных была
предложена модель - механизм полимеризации мономеров, аналогичный синтезу
ацильных цепей жирных кислот, согласно которой димеризация ПГА - синтазы
необходима для формирования активного центра с двумя идентичными тиольными
группами. Последние образуют сайты связывания с субстратом и растущей
полимерной цепью, при этом растущая цепь переходит с одной тиольной группы на
другую. Растущая цепь полимера стабилизирует димерную структуру фермента;
последний находится в клетке в гранулосвязанной форме и локализуется на
поверхности полимерных гранул.
Одним из базовых свойств
полигидроксиалканоатов является молекулярная масса. Разные микроорганизмы или
один и тот же организм в различных условиях роста синтезируют полимеры с
молекулярной массой от сотен до миллиона дальтон. Имеющиеся результаты
свидетельствуют о том, что за длину полимерной цепи отвечает ПГА-синтаза.
Впервые это было продемонстрировано в системе in
vitro при изучении
реакции полимеризации, катализируемой очищенной ПГА-синтазой из R.eutropha.
Молекулярный вес полимера снизился с увеличением концентрации фермента. Эта
закономерность была подтверждена in
vivo. Было
продемонстрировано, что увеличение экспрессии ПГА-синтазного гена из R.eutropha
приводит к снижению молекулярного веса ПГА, синтезируемого рекомбинантным
штаммом Е. coli. Косвенным
доказательством этого феномена является увеличение числа гранул в клетке при
увеличении активности ПГА-синтазы в A.Eutrophus.
Однако молекулярный вес ПГА, синтезируемый рекомбинантным штаммом R.eutropha
РНВ-4, не зависел от активности ПГА-синтазы. Поэтому авторы выдвинули
предположения о существовании цепь-переносящего фактора (chain-transfer
agent), включающегося в
регуляцию длины цепи полимера.
Синтаза, являясь ключевым
ферментом в образовании полимерной цепи, отвечает и за формирование гранул. Как
только включается синтез полимера, растворимая форма фермента переводит
мономер-КоА в олигомер, далее фермент-олигомерный комплекс образует мицеллу с
локализацией фермента на поверхности. Гидрофобные участки полимерной цепи
находятся внутри мицеллы и отделены от цитозоля. В этих условиях происходит
быстрое удлинение полимерной цепи и формируется гранула. Размер гранул зависит
от количества ПГА-синтазы в клетке, так как концентрация фермента может
определять количество зарождаемых гранул.
Рис.4 Механизм формирования
гранул ПГА.
Прежде всего, растворимая
ПГБ-синтаза взаимодействует с возрастающими концентрациями 3-оксибутирил-КоА в
цитоплазме, приводя к праймингу фермента известным механизмом. В течение
начальной лаг-фазы оксибутират олигомеры формируются медленно и вытесняются из
фермента. С увеличением длины олигомера затем формируются мицеллы.
Мицелло-подобные частицы дают границу раздела фаз с полимеразой, расположенной
внутри. Фермент затем быстро продолжает ПГБ-синтез, вытесняя большее количество
ПГБ в возрастающую гранулу.
1.4 Организация генов
биосинтеза ПГА
полигидроксиалканоат
фермент биосинтез ген
В последние годы в результате достижений в
области молекулярно-генетических исследований и детального исследования системы
синтеза полигидроксиалканоатов появились перспективы для получения
рекомбинантных штаммов как более эффективных продуцентов биополимеров. На этом
пути, ориентированном на получение высокопродуктивных биосинтетиков ПГА с
использованием разнообразных источников углерода, возможны два направления.
Первое предполагает интродукцию субстрат-утилизирующих генов в бактериальные
штаммы, синтезирующие ПГА для расширения их трофического потенциала. Второе
использует гены системы синтеза ПГА для интродукции их в быстрорастущие
микроорганизмы, не синтезирующие полигидроксиалканоаты, но характеризующиеся
широким органотрофным потенциалом. К настоящему моменту гены синтеза ПГА
клонированы из нескольких десятков бактерий, что позволило получить
разнообразные в таксономическом отношении продуценты полимеров, а также
организмы, обладающие способностью синтезировать совершенно новые типы ПГА.
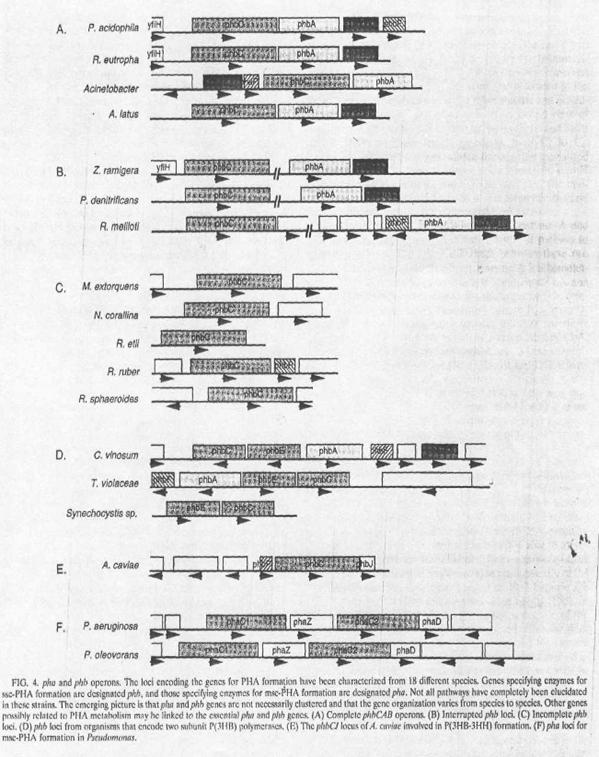
Первый phb
ген, выделенный из бактерии Z. ramigera,
был интересен для биополимерной инженерии тем, что продуцировал P(3HB) и
внеклеточный полисахарид. При использовании анти-тиолазных антител phbA
ген, несущий в Z. ramigera
генную библиотеку, был обнаружен в Escherichia
coli и впоследствии
клонирован. Установили, что phbA
и phbB гены формируют
оперон, в то время как phbC
расположен в другом месте на хромосоме Z. ramigera.
Клонирование phbA и phbB
облегчило расшифровку закодированных кетоацил-КоА - тиолазы и ацетоацил-КoA
- редуктазы, благодаря тому что выяснили кинетическую и механистическую
характеристики этих ферментов. Начиная с первоначального открытия этих phb
генов, было установлено, что многие гены кодировали ферменты, участвующие в
метаболизме ПГА, которые были клонированы из различных организмов. Учитывая
разнообразие путей биосинтеза P(3HB), не удивительно, что местоположение pha
значительно изменилось. В Acinetobacter
spp., Alcaligenes
latus, Pseudomonas
acidophila, и R. eutropha,
phbCAB гены находятся в
тандеме на хромосоме, хотя не обязательно в том же самом порядке. В Paracoccus denitrificans,
Rhizobium
meliloti, и Z. ramigera,
оперон phbAB и ген phbC
локализованы не сцепленно. ПГА-полимераза в Chromatium
vinosum, Thiocystis
violacea, и Synechocystis
- это фермент состоящий из двух субъединиц, кодирующийся phbE
и phbC генами. В этих
организмах, phbAB и phbEC
находятся в одном локусе, но ориентированы в разных направлениях. Рhb
локализован в C. vinosum,
P. acidophila, R. Eutropha,
Rhizobium
meliloti, и T. Violacea;
все они имеют дополнительный ген, phbF,
функция которого в метаболизме ПГА до настоящего времени не известна, в то
время как часть гена, кодирующая белок гипотетически гомологичный к E. сoli,
кодировала белок YfiH,
расположенный в P. acidophila,
R. eutropha, и Z. ramigera
в генах P(3HB) - полимеразы. В Methylobacterium
extorquens, Nocardia
corallina, Rhizobium
etli, Rhodococcus
ruber, и Rhodobacter
sphaeroides, был
идентифицирован к настоящему времени только ген кодирования ПГБ - полимеразы.
Ген синтеза ПГА-полимеразы в Aeromonas
caviae - это уникальная
сторона биосинтетического фермента ПГА, кодирующегося геном phaJ.
В среднецепочечном ПГА, производящегося из P. okovorans
и P. aeruginosa,оперон
pha содержит два phaC
гена отделенных от phaZ,
который кодирует внутриклеточную ПГА - деполимеразу. Две ПГА полимеразы на 50 -
60 % идентичны в их первичной структуре и имеют очень похожую специфичность к
субстрату.
Первые ПГА, выделенные из
бактерии, использовали такой путь метаболизма, который не был направлен на
воспроизведение генетической информации. Наиболее вероятно, что формирование ПГА
в этих организмах не было основным путем метаболизма, а происходило только в
результате побочных реакций.
В течение развития, phaC
иногда объединялся с генами типа phbAB
или phaJ, которые
кодируют мономер, или с генами, вовлеченными в другие аспекты метаболизма PHA,
типа phaZ. Активное
давление на выборочные гены происходит тогда, когда проведено кластеризование pha
генов в опероне в некоторых организмах (как в P.acidophila,
R.eutropha, Acinctobacter,
Alcaligenes
latus, и Aeromonas
caviae) или как отдельные
транскрипционные модули в других (как в Z. ramigera,
P. denitrificans, Rhizobium
meliloti, C, vinosum,
T. violacea, P. oleovorans,
P. putida, и возможно в
других микроорганизмах, для которых не были идентифицированы тиолазные и
редуктазные гены). Вторая эволюционная сила, должна была, воздействовать на pha
гены, так как некоторые из них, но не все, а только структурированые в
различных местоположениях, содержат phbF
и phbP гены или гомологи yfiH.
Вопрос о том, была ли наследственная ПГА - полимераза закодирована одним (phaC)
или двумя (phaEC) генами,
остается неизвестным. Начиная с систем полимеразы с двумя субъединицами, в C. vinosum
и T. violaceae
имеются гены соседней тиолазы и гены редуктазы, тогда как phaEC
в Synechocystis не производит
сшивание phaEC или
сращивания phaC, это,
возможно, предшествовало перестановкам в pha
местоположениях.
Хотя B. megaterium
был первой видом, из которого был выделен и идентифицирован P(3HB) , но его
биосинтетическая организация еще не была охарактеризована. Выделенные недавно
мутанты B. megaterium,
поврежденные в результате формирования P(3HB), позволили клонировать и
охарактеризовать phb
гены из этого исторически известного производителя P(3HB).
Несмотря на манипулирование с phb
генами в R.eutropha,
продолжался поиск более перспективного объекта для создания рекомбинантных
продуцентов ПГА. Бактерии рода Alcaligenes,
хорошо себя зарекомендовавшие в качестве промышленного продуцента ПГА и
способные с большими выходами синтезировать разнообразные по составу и
свойствам полимеры, имеют, однако, некоторые ограничения. Синтез
полигидроксиалканоатов происходит у них при низких скоростях роста, что
затрудняет процесс ферментации. Кроме этого, эти организмы не достаточно полно
охарактеризованы генетически, что ограничивает их использование для
генно-инженерного манипулирования. Этих проблем не возникает при рассмотрении
быстро растущих и наиболее полно охарактеризованных в генетическом плане
бактерий Escherichia
coli. Сконструированные
к настоящему времени рекомбинантные штаммы E.Coli,
содержащие стабильные и высококопийные плазмиды с генами синтеза
полигидроксиалканоатов из Al.eutrophus
и других бактерий, характеризуется способностью синтезировать высокие
концентрации полимеров различного состава при общей высокой продукционной способности.
Первые результаты по
клонированию pha генов Al.eutrophus
в E.Coli
дали положительные результаты, - в трансгенной бактерии зафиксировали
образование гранул ПГА.
Одной из ключевых проблем
продукции ПГА на основе трансгенных микроорганизмов является стабильность и
постоянство экспрессии phb
генов в течение ферментации. Получение полигидроксиалканоатов на основе
рекомбинантных организмов часто сопровождается снижением числа плазмиднесущих
особей в популяции, а также понижением копийности плазмид в ходе репликации.
Это может быть также следствием гибели части клеток, несущих гены устойчивости
к антибиотикам, например, parB.
Нестабильность phb
генов в высокоплотных культурах в ходе ферментации, сопровождающаяся падением
продукции ПГА, влияет на его стоимость.
Большинство генно-инженерных
штаммов, применяемых для получения ПГА, растет на глюкозе, цена которой
достаточно высока. Расширение трофического потенциала и вытекающая из этого
возможность расширения и удешевления сырьевой базы для производства ПГА чрезвычайно
значимы для расширения производства. Однако результаты, полученные в этом
направлении, пока не так значимы, как ожидалось. Так, интродукция β-галактозидазного
гена и gal оперона E.Coli
в Al.eutrophus
не позволила получить быстрорастущего на лактозе штамма. В другой работе,
полученный рекомбинантный штамм Al.eutrophus
с встроенными генами из Bacillus
subtilis, сконструированный
с целью продукции ПГА на сахарозе, также характеризовался очень медленным
ростом. Сравнительно недавно полученный рекомбинантный штамм на основе E.Coli
и K.aerogenes,
исходно утилизирующих сахарозу, способен, как, оказалось, синтезировать от 45
до 70% ПГБ при росте на среде с сахарозой (цена которой на 33-50% ниже
стоимости глюкозы). Штамм способен также включать в полимер до 55% валерата в
присутствии в среде пропионата, однако, он также не был достаточно стабильным.
В целом, полученные результаты
в области метаболической инженерии синтеза ПГА обнадеживают и позволяют
надеяться на возможность конструирования эффективных трансгенных штаммов для
получения биополимеров, обладающих:
) способностью к быстрому росту
в высокоплотных культурах с высокой общей продуктивностью;
) синтезирующих большие
количества полимеров с использованием различных углеродных субстратов, а также
) облегчить процедуру
экстракции полимера из клеточной биомассы и 4) позволяющих контролировать
внутриклеточную деполимеразную систему, деградирующую синтезированный полимер.
ЗАКЛЮЧЕНИЕ
В результате проведенной работы, был проработан
и изучен материал по теме генетика и биохимия микробного синтеза
полигидроксиалканоатов, и сделан обзор данной темы по литературным источникам.
В работе были рассмотрены физико-химические свойства ПГА, их пути синтеза,
организация генов, а также ПГА - синтаза из R.eutropha.
Способность микроорганизмов синтезировать
полигидроксиалканоаты (ПГА) различного состава вызывает большой интерес в связи
с возможностью направленного получения полимеров с заданными свойствами. Также,
способность микробных ПГА разрушаться в различных средах, представляет собой
одно из наиболее привлекательных их коммерческих свойств, поэтому является
предметом специальных исследований.
Полигидроксиалканоаты (ПГА) представляют
семейство полиэстеров, имеющих термопластические и резиновые свойства, которые
синтезируют прокариотические организмы в специфических условиях
несбалансированного роста в качестве эндогенного депо энергии и углерода,
используя для этого различные субстраты (сахара, органические кислоты, спирты и
другое).
На примере наиболее изученного из ПГА - полимера
β-оксимасляной
кислоты установлено, что пути его синтеза практически одинаковы у различных
микроорганизмов (Alkaligenes,
Azotobakter, Pseudomonas).
В биосинтезе полигидроксиалканоатов ключевыми являются три фермента: β-кетотиолаза,
ацетоацетил-КоА-редуктаза и ПГА-синтаза, которые кодируются соответственно
генами phbA, phbB,
phbC.
Первые два отвечают за синтез
субстратов, причем кетотиолаза включает синтез полимера. ПГА - синтаза
определяет количество и качество синтезированного ПГА.
В настоящее время выделено и
охарактеризовано более 50 структурных генов синтаз из разных микроорганизмов.
ПГА - синтаза из R.eutropha
состоит из одного типа субъедениц с молекулярной массой около 64 кДа и
кодируется геном phbC.
В клетке ПГА - синтаза находится в двух формах: растворимой и связанной с
гранулами полимера. Обе формы наиболее активно используются для построения
полимерных молекул короткоцепочечных (от 3 до 5 атомов углерода)
3-гидроксиацил-КоА и это свидетельствует о строгой специфичности фермента к короткоцепочечным
производным.
В рассмотрении ПГА на
молекулярно - генетическом уровне, было выяснено, что в последние годы в
результате достижений в области молекулярно-генетических исследований и
детального исследования системы синтеза полигидроксиалканоатов появились
перспективы для получения рекомбинантных штаммов как более эффективных
продуцентов биополимеров. На этом пути, ориентированном на получение
высокопродуктивных биосинтетиков ПГА с использованием разнообразных источников
углерода, возможны два направления. Первое предполагает интродукцию
субстрат-утилизирующих генов в бактериальные штаммы, синтезирующие ПГА для
расширения их трофического потенциала. Второе использует гены системы синтеза
ПГА для интродукции их в быстрорастущие микроорганизмы, не синтезирующие
полигидроксиалканоаты, но характеризующиеся широким органотрофным потенциалом.
К настоящему моменту гены синтеза ПГА клонированы из нескольких десятков
бактерий, что позволило получить разнообразные в таксономическом отношении
продуценты полимеров, а также организмы, обладающие способностью синтезировать
совершенно новые типы ПГА.
В целом, полученные результаты
в области метаболической инженерии синтеза ПГА обнадеживают и позволяют
надеяться на возможность конструирования эффективных трансгенных штаммов для
получения биополимеров, обладающих:
) способностью к быстрому росту
в высокоплотных культурах с высокой общей продуктивностью
) синтезирующих большие
количества полимеров с использованием различных углеродных субстратов, а также
) облегчить процедуру
экстракции полимера из клеточной биомассы и 4) позволяющих контролировать
внутриклеточную деполимеразную систему, деградирующую синтезированный полимер.
СПИСОК ЛИТЕРАТУРЫ
1. Волова Т.Г. Биосинтез на
водороде. - Новосибирск, 2006.-398 с.
2. Волова Т.Г., Калачёва Г.С.,
Горбунова О.В., Жила Н.О. Динамика активности ферментов клеточного цикла
полигидроксиалканоатов у Ralstoniа
eutropha//Прикладная
биохимия и микробиология. - 2008-т.40, №2-с. 201-209.
. Волова Т.Г., Войнов Н.А.
Кинетические параметры культуры водородокисляющих бактерий Ralstoniа
eutropha в режиме
биосинтеза полигидроксибутирата// Прикладная биохимия и микробиология. -
2007-т.39, №2-с. 189-193.
. Волова Т.Г., Калачёва Г.С.
Синтез сополимеров гидроксибутирата и гидроксивалерата [поли (3ГБ/3ГВ)]
бактериями Ralstoniа
eutropha// Микробиология. -
2008-т.74, №1-с. 63-69.
. Волова Т.Г., Беляева О.Г.,
Плотников В.Ф., Пузырь А.П. Исследование биодеградации микробных
полигидроксиалканоатов// Прикладная биохимия и микробиология. - 2008-т.34,
№5-с. 539-543.
6. Bernd H.A. Rehm.
Genetics and biochemistry of Polyhydroxyalkanoate granule self - assembly: The
key role of polyester synthases// Biotechnology letters, 2009, №4-p.207-213.
. Daniel K.Y.Soleiman,
Richard D. Ashby, Arland T., Hotchkiss J., Thomas A. Foglia// Byosynthesis of
medium - chain - length PHAs from soy molasses// Biotehnology letters, 2011,
№3-p.157-162.
. IL Lee, Ho Myoung
Ryu, Moo Woong Kim, Young Ha Rhee, Jeong-Yoon Kim. Identification and
heterologous expression of the PHA biosynthesis genes from Alkaligenes sp.
SH-69// Biotechnology letters, 2008,
Vol.20, №10-p.
969-975.
. Lara L. Madison, W.
Husman. Metabolic Engineering of Poly(3- Hydroxyalkanoates): From DNA to
Plastic// Microbiology and Molecular Biology Reviews, Mar.2009, p.21-53.
. Yahaya M.Normi,
Tomohiro Hiraishi, Seiichi Taguchi, Kunar Sudesh? Nazalan Najimudin, Yoshihary
Doi. Site - directed saturation mutagenesis at residue F420 and recombinations
with another beneficial mutation of Ralstonia eutropha PHA synthase//
Biotechnology letters, 2006, №27-p. 705-712.
. Doi Y., Kanesawa Y.,
Kunioka M., Yamada K., Biodegradation of biosynthetic and chemosynthetic
polyhydroxyalkanoates. In: Doi Y., Fukuda K., (eds) Biodegradable polymers and
plastics, Elsevier, Amsterdam, 2006, p. 39-51.
Summary
(PHAs) are a family of polyesters
produced in bacteria as a carbon and energy reserve. Depending on their
chemical structure, PHAs can have properties ranging from stiff and brittle plastics
to elastomers and rubbers. These polymers are completely biodegradable in the
environment, being metabolized by microorganisms to carbon dioxide and
water.are accumulated as discrete granules to levels as high as 90% of the cell
dry weight and are generally believed to play a role as sink for carbon and
reducing equivalents. When nutrient supplies are imbalanced, it is advantageous
for bacteria to store excess nutrients intracellularly, especially as their
general fitness is not affected. By polymerizing soluble intermediates into
insoluble molecules, the cell does not undergo alterations of its osmotic state
and leakage of these valuable compounds out of the cell is prevented.
Consequently, the nutrient stores will remain available at a relatively low
maintenance cost and with a secured return on investment.PHAs are extracted
from the bacterial cell, however, these molecules show material properties that
are similar to some common plastics such as polypropylene. The bacterial origin
of the PHAs makes these polyesters a natural material, and, indeed, many
microorganisms have evolved the ability to degrade these macromolecules.
Besides being biodegradable, PHAs are recyclable like the petrochemical
thermoplasts. This review summarizes the chemical and physical properties of
PHAs and the biochemical and genetic studies of the pathways involved in PHA
metabolism. Within this framework, the scientific advances that have been made
with the available pha genes for economic PHA production processes will be described.