Трубчатые кости
Алгебри
інваріантності та точні розв’язки математичної моделі хемотаксису
Зміст
Вступ
Розділ 1. Застосування систем
рівнянь хемотаксису в математичній біології
Розділ 2. Симетрії Лі однієї
системи рівнянь хемотаксису
.1 Виведення системи визначальних
рівнянь
.2 Розв’язування системи
визначальних рівнянь
.3 Аналіз отриманих алгебр
інваріантності
Розділ 3. Редукція системи ДРЧП
до системи ЗДР
Висновки
Список літератури
Вступ
На сьогоднішній день вже
загальноприйнятою є теза про те, що переважна більшість реальних процесів, які
вивчаються в фізиці, біології, хімії тощо, мають суттєво нелінійну природу, а
тому математично описуються нелінійними рівняннями. У переважній більшості - це
нелінійні диференціальні рівняння з частинними похідними (ДРЧП) та системи
(ДРЧП). Тому побудова адекватних моделей цих процесів у формі деяких класів
нелінійних ДРЧП та розвиток нових підходів до розв'язання цих рівнянь є дуже
актуальною проблемою.
Як відомо, проблема розвитку нових
підходів для інтегрування відомих нелінійних ДРЧП з тривіальною (найпростішою)
симетрією Лі, які є базовими для реальних процесів, є надзвичайно актуальною. В
90-х роках різними дослідниками, зокрема, П. Олвером, В.А. Галактіоновим, С.Р.
Свірщевським, А.С. Фокашем, К.М. Ліу, В.І. Фущичем, Р.М. Чернігою, М.К. Нучі
запропоновано декілька нових підходів для побудови точних розв'язків нелінійних
рівнянь. Хоч жоден з цих підходів не можна розглядати як абсолютно новий
універсальний метод інтегрування нелінійних ДРЧП, проте їхнє застосування до
розв'язання низки відомих нелінійних рівнянь дозволило побудувати нові
розв'язки, які неможливо знайти не тільки методом Лі, а часто - і методом
некласичних симетрій Блумана - Коула.
Метод Лі названий за іменем його
фундатора - норвезького математика Софуса Лі, має більш широкі межі
застосування, оскільки він застосовний і до неінтегровних нелінійних рівнянь з
частинними похідними. Метод ґрунтується на знаходженні та застосуванні
операторів алгебри інваріантності (симетрій Лі) розглядуваного нелінійного рівняння
з частинними похідними для знаходження його точних розв'язків. Хоч базові
теореми цього методу сформульовані ще С.Лі, одначе протягом останніх сорока
років цей метод невпинно розвивається і регулярно з'являються роботи, в яких
автори одержують нові результати для нелінійних рівнянь з нетривіальною
симетрією Лі. Найбільший внесок у розвиток та застосування методу Лі за цей час
зробили Л.В. Овсянніков, Дж. Блуман, П. Олвер, Н.Х. Ібрагімов, В.І. Фущич. В
Україні перші роботи на цю тему були опубліковані львівським математиком В.Г.
Костенком наприкінці 50-х років.
У природі усі живі істоти прагнуть
опинитися в областях з найбільш сприятливими для їх виду умовами. Це твердження
справедливо не лише для багатоклітинних організмів, але і для одноклітинних, і
навіть для окремих клітин. Добре відомо, що ендотеліальні клітини капілярної
мережі, макрофаги і цілий ряд інших клітин мають хемотактичну рухливість,
тобто, здатні змінювати напрям свого руху у відповідь на зовнішній сигнал.
Зазвичай таким сигналом є розчинена хімічна речовина, яка називається
хемоатрактантом, проте існує і інший тип спрямованого руху, гаптотаксис - рух
по градієнту напруги міжклітинного матриксу. Окрім руху по градієнту
аттрактанта, експериментально спостерігається зміна швидкості випадкового руху
клітин залежно від концентрації хімічного медіатора, так званий хемокінез.
Хемотаксис і хемокінез відіграють важливу роль в процесах формоутворення
(морфогенезу), імунних і регуляторних процесах. Крім того, вважається, що
багато типів трансформованих клітин, особливо метастатичний активних, володіють
хемотактичною рухливістю і хемокінезом.
Однією з перших робіт по
математичному моделюванню, що враховує хемотаксис, була робота Патлака,
опублікована в 1953 році , яку потім розвинули Келлєр і Зегель в 1970 .
Використаний ними математичний запис хемотактичного члена став загальновизнаним
для опису направленного руху клітин і одноклітинних організмів в різних
біологічних явищах.
Існує ряд робіт по математичному
моделюванню росту пухлини, в яких враховується хемотактична і хаптотактична
рухливість пухлинних клітин . Проте коректне врахування хемотактичної
рухливості украй ускладнений через відсутність скільки-небудь достовірних
експериментальних даних відносно характерної величини цієї рухливості в тканині.
Дійсно, практично усе дослідження хемотаксису і хемокінезу пухлинних клітин
проводяться in vitro( іn vitro - це техніка виконання експерименту у пробірці,
або, більш загально, у контрольованому середовищі поза живим організмом) в
камері Бойдена або за допомогою спеціальних плоских дисперсійних поверхонь.
Проте ці дослідження дозволяють порівнювати величини хемотактичної рухливості у
різних клітинних ліній або ж стежити за її зміною з плином часу або залежно від
градієнта хімічного медіатора (хемоатрактанту). Відсутність кількісних даних
про величину хемотактичної рухливості в щільній тканині робить її врахування в
математичній моделі досить спекулятивним. Дійсно, завжди можна підібрати
параметри хемотактичного члена так, щоб в результаті отримати необхідний ефект.
Розповсюдження бактеріальних
популяційних хвиль добре описується математичними моделями, основаними на
рівняннях Келлєра-Зегеля
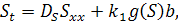
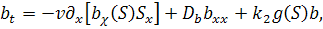
де 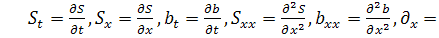 причому
причому 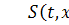 концентрація субстрату-аттрактанту,
який споживається бактеріями,
концентрація субстрату-аттрактанту,
який споживається бактеріями, 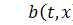 концентрація бактерій,
концентрація бактерій, 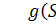 питома швидкість росту бактерій,
питома швидкість росту бактерій, 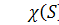 функції хемотаксисної відповіді,
функції хемотаксисної відповіді, 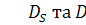 коефіцієнти дифузії субстрату та
бактерій відповідно;
коефіцієнти дифузії субстрату та
бактерій відповідно; 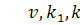 сталі;
сталі; 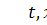 часова та просторова змінні
відповідно. Моделлю Келлєра-Зегеля та її деякими модифікаціями описується також
формування та поширення хемотаксисних кілець Адлера та різні процеси
структуроутворення в бактеріальних колоніях при їх взаємодії.
часова та просторова змінні
відповідно. Моделлю Келлєра-Зегеля та її деякими модифікаціями описується також
формування та поширення хемотаксисних кілець Адлера та різні процеси
структуроутворення в бактеріальних колоніях при їх взаємодії.
Метою магістерської роботи є
дослідження узагальненої системи Келлєра - Зегеля теоретико - алгебраїчними
методами та методами якісної теорії диференціальних рівнянь.
Актуальність роботи. Класична
модель Келлєра - Зегеля - система нелінійних диференціальних рівнянь з
частинними похідними - є базовою моделлю для опису значної кількості
біологічних процесів, для яких характерний спрямований рух . Симетрійні
властивості такої системи та її узагальнень, а також побудова точних розв'язків
ще недостатньо вивчені.
Предметом вивчення є алгебри
інваріантності та метод анзаців і їх застосування до побудови точних розв'язків
нелінійних рівнянь.
Об'єкт вивчення - диференціальні
рівняння з частинними похідними другого порядку, які є математичними моделями
процесу хемотаксису.
Гіпотеза дослідження -
оператори інваріантності системи Келлєра - Зегеля дозволяють побудувати класи
точних розв'язків цієї системи.
Методи дослідження -
теоретико - алгебраїчні методи та методи якісної теорії диференціальних
рівнянь.
Магістерська робота
складається зі вступу, трьох розділів, висновків, додатку та списку
використаної літератури.
У вступі формулюється мета
роботи, актуальність теми, об'єкт, гіпотеза та методи дослідження.
Перший розділ
характеризується описанням значення процесу хемотаксису в математичній біології
і в житті різних організмів в цілому.
У другому розділі виведена
система визначальних рівнянь, подано подане розв'язання цієї системи, а також
проаналізовані алгебри інваріантності, які були утворені при розв'язанні
системи визначальних рівнянь, зокрема, побудовані базиси отриманих алгебр
інваріантності і обчислені комутатори.
Третій розділ присвячено
побудові анзаців і проведенню редукції системи диференціальних рівнянь з
частинними похідними до системи звичайних диференціальних рівнянь.
У висновках коротко
висвітлено основні положення роботи.
В списку літератури
наводиться перелік основних літературних джерел, які використовувалися при
написанні магістерської роботи.
Розділ 1. Застосування систем
рівнянь хемотаксису в математичній біології
Сучасна наука
характеризується глибоким проникненням математичних методів у її різні галузі.
З середини 50-х років у всьому світі почався стійкий ріст інтересу до
використання математики в екології та біології. Математичне моделювання
допомагає створити цілісну картину того чи іншого явища, передбачити різні
наслідки певного порушення функціонування екосистеми в локальному або глобальному
масштабі .
Впродовж останніх десятиліть
намітився значний прогрес в кількісному (математичному) описі функцій різних
біосистем на різних рівнях організації життя : молекулярному, клітинному,
органному, організменому, популяційному, біогеоценогенному (экосистемному).
Життя визначається безліччю різних характеристик цих біосистем і процесів, що
протікають на відповідних рівнях організації системи і інтегрованих в єдине
ціле в процесі функціонування системи. Про моделі, що базуються на істотних
постулатах про принципи функціонування системи, які описують і пояснюють
широкий круг явищ і виражають знання в компактній, формалізованій формі, можна
говорити як про теорію біосистеми.
Побудова математичних моделей
(математичних теорій) біологічних систем стала можливою завдяки виключно
інтенсивній аналітичній роботі експериментаторів : морфологів, біохіміків,
фізіологів, фахівців з молекулярної біології та ін. В результаті цієї роботи
кристалізовані морфофункціональні схеми різних клітин, у рамках яких
впорядковано у просторі та часі протікають різні фізико-хімічні і біохімічні
процеси, що утворюють дуже складне переплетення. Другою, дуже важливою
обставиною, яка сприяє залученню математичного апарату в біологію, є ретельне
експериментальне визначення констант швидкостей багатьох внутрішньоклітинних
реакцій, що визначають функції клітини і відповідної біосистеми. Без знання
таких констант неможливий формально-математичний опис внутрішньоклітинних
процесів. І, нарешті, третьою умовою, що визначила успіх математичного моделювання
в біології, є розвиток потужних обчислювальних засобів у вигляді персональних
комп'ютерів і суперкомп'ютерів. Це пов'язано з тим, що зазвичай процеси,
контролюючи ту або іншу функцію клітин або органів, численні, охоплені петлями
прямого і зворотного зв'язку і, отже, описуються системами нелінійних рівнянь.
Такі рівняння не розв'язуються аналітично, але можуть бути розв'язані чисельно
за допомогою комп'ютера. Чисельні експерименти на моделях, здатні відтворювати
широкий клас явищ в клітинах, органах і організмі, дозволяють оцінити
правильність припущень, зроблених при побудові моделей. Зауважимо, що, хоча в
якості постулатів моделей використовуються експериментальні факти, необхідність
деяких допущень і припущень є важливим теоретичним компонентом моделювання. Ці
допущення і припущення є гіпотезами, які можуть бути піддані експериментальній
перевірці. Таким чином, моделі стають джерелами гіпотез, і притому,
експериментально верифікуючих. Експеримент, спрямований на перевірку цієї
гіпотези, може спростувати або підтвердити її і тим самим сприяти уточненню
моделі. Така взаємодія моделювання і експерименту відбувається безперервно,
призводячи до усе більш глибокого і точного розуміння явища : експеримент
уточнює модель, нова модель висуває нові гіпотези, експеримент уточнює нову
модель і так далі.
Нині математична біологія, що
включає математичні теорії різноманітних біологічних систем і процесів,
являється, з одного боку, вже науковою дисципліною, що достатньою мірою
склалася, а з іншого боку, однією з наукових дисциплін, що найбурхливіше
розвиваються, об'єднують зусилля фахівців з різних областей знання -
математиків, біологів, фізиків, хіміків і фахівців з комп'ютерних наук. Можна з
упевненістю говорити про ряд дисциплін математичної біології, що найбільш
сформувалися : математичну генетику, імунологію, епідеміологію, екологію, ряд
розділів математичної фізіології, зокрема, про математичну фізіологію
серцево-судинної системи. Як всяка наукова дисципліна, математична біологія має
свої предмет, способи, методи і процедури дослідження. В якості предмета
досліджень виникають математичні (комп'ютерні) моделі біологічних процесів, що
одночасно є і об'єктом дослідження, і інструментом для дослідження власне
біологічних систем. У зв'язку з такою двоякою суттю біометричних моделей, вони
мають на увазі використання наявних і розробку нових способів аналізу
математичних об'єктів (теорій і методів відповідних розділів математики) з
метою вивчення властивостей самої моделі як математичного об'єкту, а також
використання моделі для відтворення і аналізу експериментальних даних, що
отримуються в біологічних експериментах. При цьому в якості одного з найбільш
важливих призначень математичних моделей (і математичної біології в цілому)
являється можливість передбачення біологічних явищ і сценаріїв поведінки
біосистеми в певних умовах і їх теоретичного обгрунтування до (або навіть
замість) проведення відповідних біологічних експериментів. Основним методом
дослідження і використання складних моделей біологічних систем є обчислювальний
комп'ютерний експеримент, який вимагає застосування адекватних методів
обчислень для відповідних математичних систем, алгоритмів обчислень, технологій
розробки і реалізації комп'ютерних програм, зберігання і обробки результатів
комп'ютерного моделювання. Ці вимоги мають на увазі розробку загальних і
спеціальних теорій, методів, алгоритмів і технологій комп'ютерного моделювання
у рамках різних напрямів біоматематики, що саме по собі є складовою частиною
наукової дисципліни "Математична біологія".
Нарешті, у зв'язку з основною
метою використання біометричних моделей для пізнання законів функціонування
біологічних систем, усі стадії розробки і використання математичних моделей
припускають обов'язкову опору на теорію і практику біологічної науки.
Хемотаксисом називають рух
мікроорганізмів, рослин і тварин, а також рухливих клітин (лейкоцитів) під
впливом хімічних речовин. Оскільки однією з важливих ознак організму є його
здатність рухатися і обмінюватися речовиною з довкіллям, без перебільшення
можна сказати, що до хемотаксису в тій або іншій мірі схильні усі живі форми.
Незважаючи на малі розміри
(близько 1 мкм), бактерійна клітина має дуже складну морфологічну структуру.
ДНК бактерії E. coli містить 5 тис. генів, з яких сто відповідальні за
рухливість і хемотаксис. Це означає, що в будівництві клітини і її
функціонуванні беруть участь тисячі білків, близько ста з яких формують руховий
і хемочуттєвий апарат. Системні зв'язки між структурами простої бактерії такі
складні, що не піддаються найпотужнішому комп'ютерному аналізу. Проте основні
закономірності руху бактерій до теперішнього часу вивчені досить детально.
За останні три десятиліття
дослідження хемотаксису мікроорганізмів вилилося в самостійну область біології
прокаріот, що добилася значних успіхів як в розумінні біофізичних і
молекулярно-біохімічних процесів, що лежать в основі бактерійної рухливості і
хемотаксису, так і в розвитку специфічних прийомів і методів дослідження.
Вже Левенгук відкрив
здатність бактерій скупчуватися навколо шматочків їжі. Проте цілеспрямовані
дослідження хемотаксису були розпочаті лише у кінці XIX століття. Зокрема,
Пфеффер запропонував простий кількісний спосіб виміру хемотаксису. У його
роботах були закладені основи термінології і уперше використана класична
методика підрахунку числа бактерій, що входять в капілярну трубку, заповнену
розчином досліджуваної речовини.
Хімічні речовини, які
притягають мікроорганізми, дістали назву аттрактантів, а відразливі -
репелентів. Саме ж явище рухової реакції мікроорганізмів на хімічний подразник
отримала назва "хемотаксис". Найкращими аттрактантами і репелентами
виявилися органічні речовини: цукор, амінокислоти, спирти. Крім того, характер
хемотаксичної реакції сильно залежав від природи випробовуваної хімічної
сполуки і виду бактерій. Дослідження Рочерта показали, що рух на аттрактанти
можна вибірково пригнічувати, інгібіруючи реакцію на м'ясний екстракт
підвищенням концентрації етилового спирту або хлороформу.
Дослідження по хемотаксису,
зроблені у кінці XIX століття, носили розрізнений характер і не привели до
створення широкого напряму досліджень, що, мабуть, було пов'язано з відсутністю
у той час серйозної техніки і наукової основи, що дозволила б досягти певного
успіху в розумінні явища. Подальші дослідження хемотаксису фактично припинилися
аж до 60-х років XX століття.
На початку 60-х років
проблемою хемотаксису зацікавився Адлер. В якості основного об'єкту він обрав
кишкову паличку E. coli, для якої структурні особливості і біохімічні процеси
були найдетальніше вивчені. Він застосував і удосконалив метод капіляра, раніше
використаний Пфеффером. Адлер уперше довів наявність у бактерій специфічних
білків-рецепторів, що пізнають певні хімічні речовини - аттрактанти і
репеленти. Це послужило поштовхом до серії біохімічних і генетичних досліджень
природи бактерійних хеморецепторів, виконаних на початку 70-х років XX століття
головним чином в лабораторіях Адлера і Кошланда.
У 1972 році Берг і Браун
сконструювали мікроскоп, що автоматично стежить за рухом окремих бактерій.
Генетичні і біохімічні дані по хемотаксису істотно доповнилися детальними
відомостями про характер руху бактерій. Було показано, що за відсутності
просторових або тимчасових змін концентрацій аттрактантів і репелентів бактерії
рухаються по гладких траєкторіях (зі швидкостями приблизно 10-20 мкм/с),
зупиняючись і перекидаючись приблизно через рівні проміжки часу (~ 1 с). Після
перекидання (що триває близько 0,1 с) бактерія починає плисти в напрямі, не
залежному від попереднього. Загалом картина руху бактерії нагадує броунівський
рух часток - випадкові блукання з деякою середньою довжиною пробігу.
Екологія мікроорганізмів,
моніторинг довкілля, біосенсорика - ось далеко не повний перелік дисциплін, де
хемотаксис бактерій міг би знайти застосування. Річ у тому, що хемотаксична система
бактерій дуже чутлива до щонайменших змін життєво важливих хімічних компонентів
довкілля і поріг хемосенсорної чутливості складає мільйонні і навіть мільярдні
долі молярної концентрації таких речовин. Недаремно хемотаксична система
бактерій сприяла і сприяє виживанню видів в ході еволюції. Як і будь-яка інша
життєво важлива функція, система хемотаксису бактерій не існує сама по собі,
незалежно від інших функцій організму. Встановлений тісний зв'язок системи
хемотаксису з дихальною системою клітини, з процесами метилування-деметилування
трансмембранних білків (МСР-білків), з фосфотрансферазною системою ФТС
(відповідальною за енергетичні процеси, пов'язані з перенесенням фосфатної
групи), з рівнем внутрішньоклітинного pH, циклічного гуанозинмонофосфата
(цГМФ).
Відомо, що забруднення
довкілля поліциклічними ароматичними вуглеводнями (ПАУ) у вигляді промислових
викидів підприємств хімічної і нафтохімічної промисловості є серйозною загрозою
здоров'ю населення і природі. Виявляється, біодеградація ПАУ мікроорганізмами
йде інтенсивніше в ризосферній сфері рослин, що примикає до кореня, чим
безпосередньо в грунті. Самі ж мікроорганізми скупчуються у коренів рослин
завдяки хемотаксису. Знання законів хемотаксису бактерій може допомогти у
вирішенні важливих питань утилізації промислових відходів.
На закінчення коротко
зупинимося на порушенні хемочуттєвості під дією деяких хімічних речовин. Окрім
згаданих етанолу і хлороформу хемотаксис блокують бензол, ацетон, багато видів
антибіотиків, а також різні речовини наркотичної дії, анестетики, адреналін,
ацетілхолін. Механізм їх дії дуже різноманітний. Швидкість розщеплювання
бактеріями деяких органічних сполук також корелює з їх хемотаксичною здатністю.
Наприклад, морські бактерії
активно розщеплюють альбумін і казеїн. Проте дія мікроскопічними концентраціями
вуглеводнів, толуолу, фенолу, відходами нафти, 2-4-дихлорфеноксиацетата,
О-О-дихлорбифенілу сильно інгібірує цей процес. В той же час такі малі
концентрації практично не чинять прямої згубної дії на життєдіяльність
мікрофлори. Основний ефект їх дії, мабуть, зводиться до пригнічення
хеморецепторної активності.
Таким чином, різноманітні
забруднення прісноводих і морських водойм навіть в малих концентраціях можуть
привести до серйозних порушень екологічного балансу. При таких малих
концентраціях це в першу чергу пов'язано з порушеннями в роботі хемосенсорних
систем мікроорганізмів. Виникла у зв'язку з цим нова область вивчення
мікробіологічних співтовариств останніми роками отримує інтенсивний розвиток.
Дослідження в живій природі доповнюються різноманітними лабораторними
експериментами.
Хемотаксис відіграє важливу
роль в процесах морфогенезу і структуроутворення в живій природі. Цю
властивість мають як одноклітинні організми, так і окремі клітини
багатоклітинних організмів. Експерименти in vitro показують, що багато типів
пухлинних клітин, особливо метастатично активні, також мають хемотаксис. Існує
цілий ряд робіт по моделюванню росту і інвазії пухлини, що використовують
модель Келлєра - Зегеля для врахування хемотаксису. Ракові клітини продукують
хімічні сполуки (реагенти), які суттєво впливають на подальший ріст та рух
ракових клітин.
В свою чергу можна ввести
хімічні сполуки, які будуть сповільнювати цей процес (хіміотерапія).
Отже, є позитивна та
негативна хемотаксична реакція. Хемотаксис називається позитивним, якщо клітина
рухається у напрямку збільшення концентрації хімічного сигналу, та негативним,
якщо вона рухається у протилежному напрямку. Для визначення розглянемо
позитивну реакцію.
Нехай 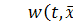 концентрація хімічної сполуки, яка
викликає хемотаксис щодо клітин,
концентрація хімічної сполуки, яка
викликає хемотаксис щодо клітин, 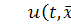 концентрація біоклітин, організмів,
концентрація біоклітин, організмів,
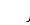 вектор
потоку біоклітин.
вектор
потоку біоклітин.
Закон збереження має вигляд:
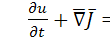 (1.1.1)
(1.1.1)
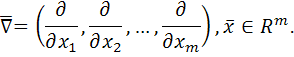
Для опису кінетики
біологічних процесів не існує достатніх загальних закономірностей, які б
дозволяли написати аналог (1.1.1).
Кінетика біологічного руху
задається формальним введенням деякої функції  у праву частину (1.1.1)
у праву частину (1.1.1)
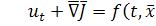 (1.1.2)
(1.1.2)
Вектор потоку біоклітин буде
рівний
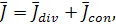
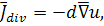
де  коефіцієнт дифузії,
коефіцієнт дифузії, 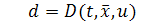 ,
,
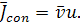
Підставимо дані вирази в
(1.1.2):
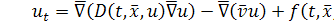 (1.1.3)
(1.1.3)
Формула (1.1.3) зображає
загальний закон біологічного руху для популяцій організмів чи біологічних
клітин.
Експериментально встановлено,
що потік, викликаний хемотаксисом, описується формулою
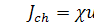 ,
,
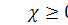 хемотаксична чутливість (
хемотаксична чутливість ( 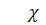 при негативному хемотаксичному
русі) .
при негативному хемотаксичному
русі) .
Вектор потоку біоклітин буде
рівний
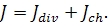
Тоді
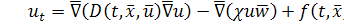 (1.1.4)
(1.1.4)
Тепер ми повинні ще записати
рівняння для еволюції хімічної сполуки, тобто закон для хімічного інгредієнта:
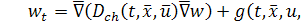 (1.1.5)
(1.1.5)
де 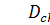 коефіцієнт дифузії хімічного
інгредієнта, функція
коефіцієнт дифузії хімічного
інгредієнта, функція  задає кінетику хімічного
інгредієнта
задає кінетику хімічного
інгредієнта  .
.
Рівняння (1.1.4) - (1.1.5)
називаються загальною системою рівнянь для опису процесу хемотаксису.
Класичним прикладом системи
(1.1.4) - (1.1.5) є модель Келлєра - Зегеля для опису реакції бактерій на
хімічний реагент:
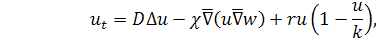
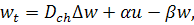
де 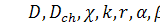 задані невідомі параметри,
задані невідомі параметри, 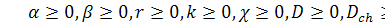 .
.
Сформулюємо крайову задачу
для випадку 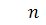 з нульовими умовами Ноймана для
системи Келлєра - Зегеля в області
з нульовими умовами Ноймана для
системи Келлєра - Зегеля в області
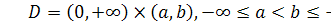
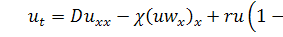 (1.1.6)
(1.1.6)
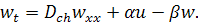
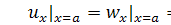 (1.1.7)
(1.1.7)
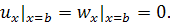
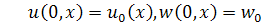 (1.1.8)
(1.1.8)
де 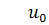 та
та 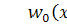 відомі гладкі функції, які задають
розподіл в початковий момент часу
відомі гладкі функції, які задають
розподіл в початковий момент часу 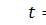
У випадку, коли коефіцієнти
дифузії дуже малі, тобто 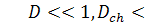 крайова задача (1.1.6) - (1.1.8)
може бути зведеною до задачі:
крайова задача (1.1.6) - (1.1.8)
може бути зведеною до задачі:
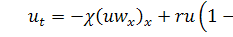 (1.1.9)
(1.1.9)
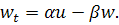
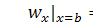 (1.1.10)
(1.1.10)
та початкових умов (1.1.8).
Задача (1.1.8) - (1.1.10) є
значно простішою, оскільки друге рівняння в (1.1.9) є лінійним звичайним
диференціальним рівнянням, що містить змінну як параметр.
Оскільки стаціонарні точки 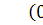 та
та 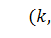 є вузлами, причому перша -
нестійкий, а друга - стійкий, то по аналогії з диузивною системою Лотки -
Вольтера важливо дослідити властивості розв'язків типу біжучої хвилі для
системи Келлєра - Зегеля (1.1.6).
є вузлами, причому перша -
нестійкий, а друга - стійкий, то по аналогії з диузивною системою Лотки -
Вольтера важливо дослідити властивості розв'язків типу біжучої хвилі для
системи Келлєра - Зегеля (1.1.6).
З цією метою систему (1.1.6)
спочатку зводять до безрозмірного вигляду за допомогою підстановки:

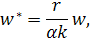
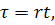
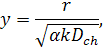
де 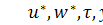 нові безрозмірні змінні.
нові безрозмірні змінні.
У підсумку отримаємо систему:
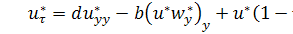 (1.1.11)
(1.1.11)
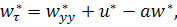
де 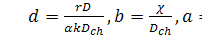
Система (1.1.11) є моделлю
Келлєра - Зегеля в безрозміреному вигляді.
Розділ 2. Симетрії Лі однієї
системи рівнянь хемотаксису
§ 2.1 Виведення системи
визначальних рівнянь
Розглянемо систему
диференціальних рівнянь з частинними похідними другого порядку, яка описує
процес хемотаксису:
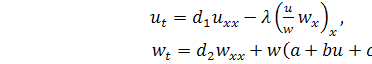
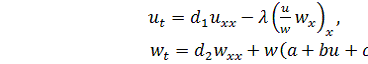 (2.1.1)
(2.1.1)
де 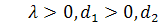 ,
,  щільність клітин,
щільність клітин, 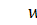 концентрація хімічної сполуки, яка
викликає хемотаксис,
концентрація хімічної сполуки, яка
викликає хемотаксис, 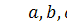 довільні дійсні числа.
довільні дійсні числа.
Перепозначивши в системі
(2.1.1) змінні 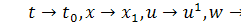 і взявши похідну в першому
рівнянні,отримаємо систему в нових позначеннях :
і взявши похідну в першому
рівнянні,отримаємо систему в нових позначеннях :
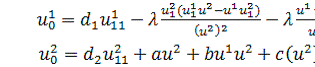 (2.1.2)
(2.1.2)
Подіємо на систему (2.1.2)
другим продовженням інфінітезимального оператора
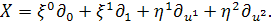
Розглянемо спочатку друге
рівняння системи:
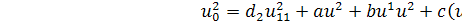 (2.1.3)
(2.1.3)
Подіявши оператором
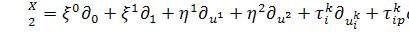
на рівняння (2.1.3) і
використовуючи формули продовження координати інфінітезимального оператора для  і
і  , а саме
, а саме
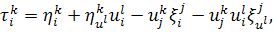
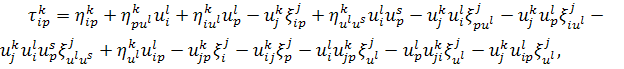
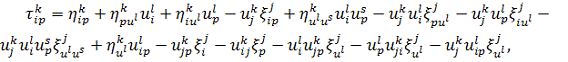
одержимо

Будемо вважати змінні 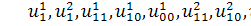 вільними змінними, а
вільними змінними, а  та
та 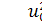 залежними. Оскільки в систему
(2.1.2) змінна
залежними. Оскільки в систему
(2.1.2) змінна  не входить, то розщепимо дане
рівняння по . Будемо мати
не входить, то розщепимо дане
рівняння по . Будемо мати
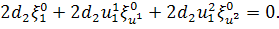
Так як  ,то на число
,то на число  можна скоротити. Аналогічно
розщепимо отримане рівняння по
можна скоротити. Аналогічно
розщепимо отримане рівняння по 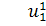 . Звідси отримаємо,що
. Звідси отримаємо,що
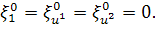
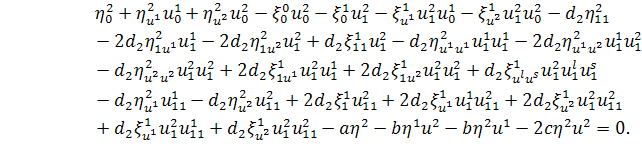
Перейдемо на многовид,
заданий системою (2.1.2), тобто замість 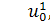 підставимо їх значення
підставимо їх значення
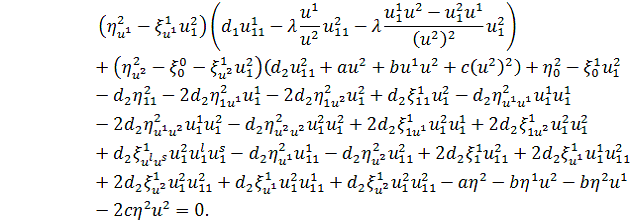
Отримане рівняння розщепимо
по 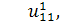 . Будемо мати наступну систему
. Будемо мати наступну систему
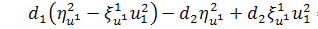 ,
,
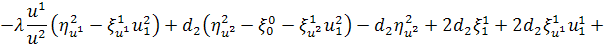
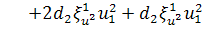 ,
,
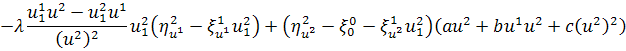
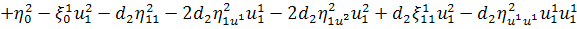
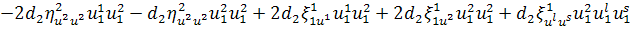
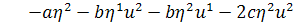 .
.
Перше рівняння з системи
розщепимо по  :
:
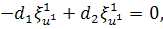
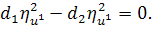
Звідси маємо, що 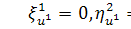
Аналогічно розщепимо друге
рівняння системи по
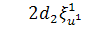 ,
,
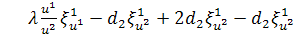 ,
,
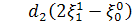 .
.
Отримаємо, що 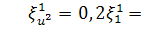
З третього рівняння системи
при умові, що 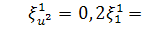 будемо мати
будемо мати
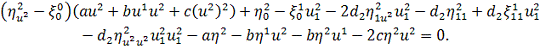
Розщепимо останнє рівняння по
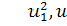 :
:
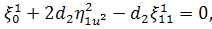

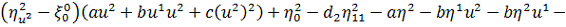
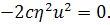
Продиференціюємо рівняння по  :
:
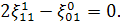
Так як 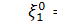 то звідси випливає, що
то звідси випливає, що 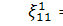
Остаточно отримаємо наступну
систему визначальних рівнянь для другого рівняння системи (2.1.2):
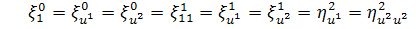 ,
,
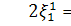 ,
,
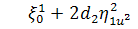 , (2.1.4)
, (2.1.4)
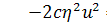 .
.
Розглянемо зараз перше
рівняння системи (2.1.2):
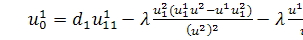 тобто
тобто
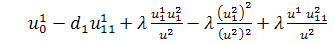 .
.
Виконуючи аналогічну
процедуру, будемо мати
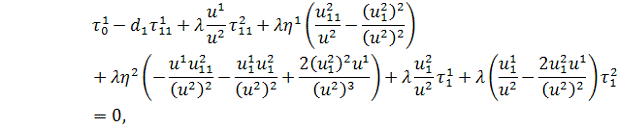
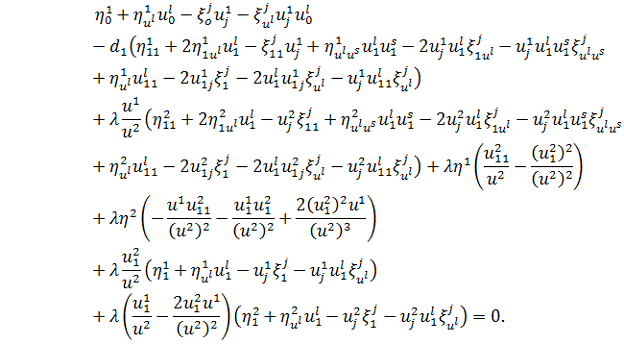
Використовуючи умови з попереднього
рівняння, отримаємо:
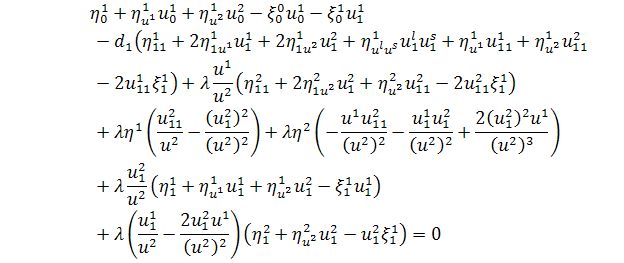
Замість підставимо їх значення
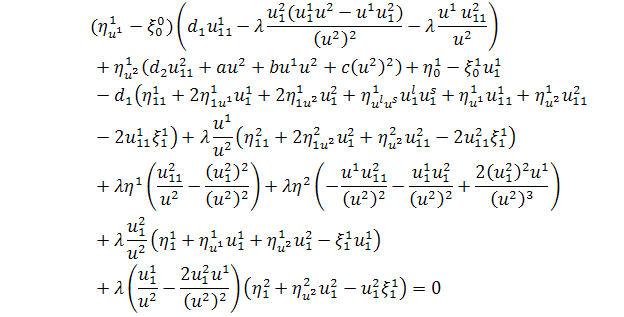
Розщепимо дане рівняння по . Будемо мати наступну систему:
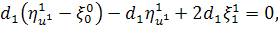
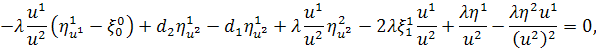
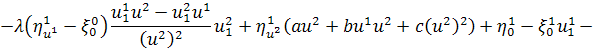
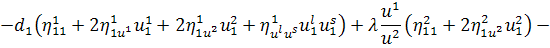
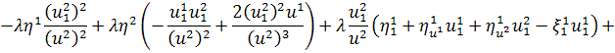
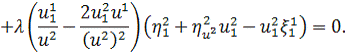
Спростивши останню систему,
отримаємо:
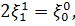
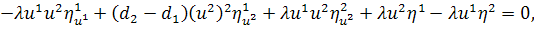
Розщепимо третє рівняння
останньої системи по 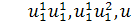
,
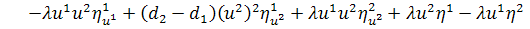 ,
,
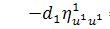 ,
,
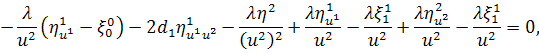
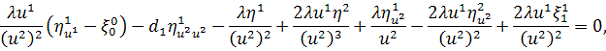
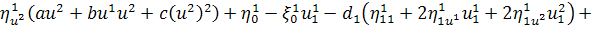
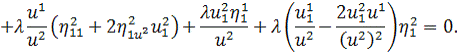
Останнє рівняння системи
розщепимо по
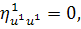
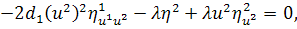
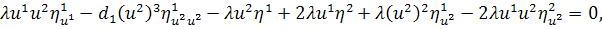

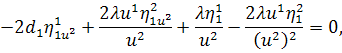
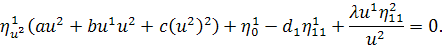
Враховуючи останню систему і систему
(2.1.4), будемо мати
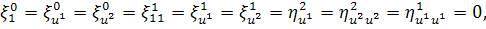
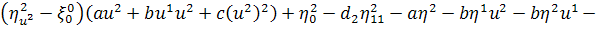
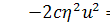 (2.1.5)
(2.1.5)
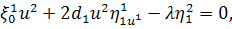
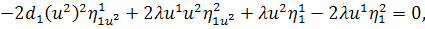
Остання система і є системою
визначальних рівнянь системи (2.1.2).
§2.2 Розв'язання системи
визначальних рівнянь
Оскільки 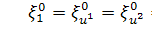 , то функція залежить лише від змінної, тобто
, то функція залежить лише від змінної, тобто 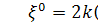 . Так як
. Так як 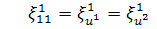 і , то функція має вигляд
і , то функція має вигляд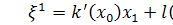 . Функція виражається рівністю
. Функція виражається рівністю 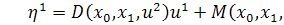 , бо
, бо 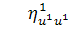 . Для функції справедливо, що
. Для функції справедливо, що 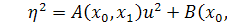 , так як
, так як 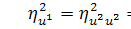
Отже, для координат
інфінітезимального оператора маємо наступні співвідношення:
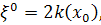
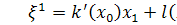 (2.2.1)
(2.2.1)
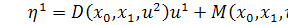
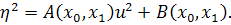
Візьмемо похідні від функцій 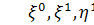 по змінних
по змінних 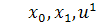 першого і другого порядку і
підставимо у систему визначальних рівнянь:
першого і другого порядку і
підставимо у систему визначальних рівнянь:
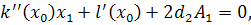
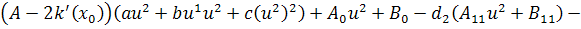
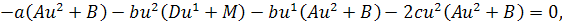
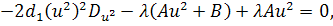

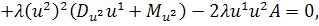

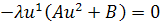
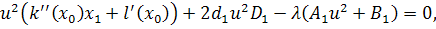
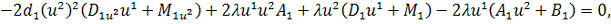
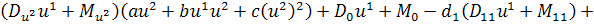
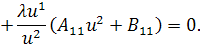
Так як функції 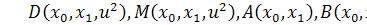 не залежать від змінної
не залежать від змінної  , то розщепимо рівняння останньої
системи по цій змінній
, то розщепимо рівняння останньої
системи по цій змінній
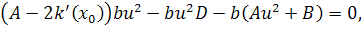
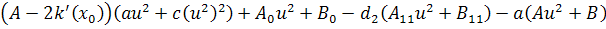

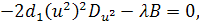
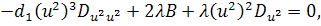
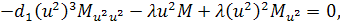
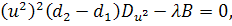
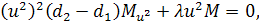
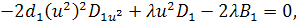
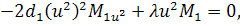
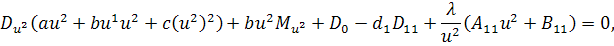
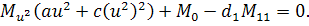
Від восьмого рівняння в
останній системі віднімемо п'яте рівняння, внаслідок чого будемо мати, що 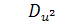 , а звідси випливає, що
, а звідси випливає, що 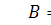
Тоді отримаємо систему:
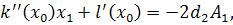
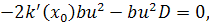
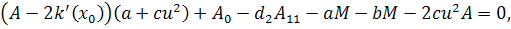

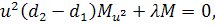
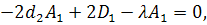
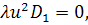
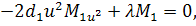
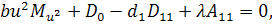
З сьомого рівняння отриманої
системи бачимо, що 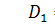 .
.
Продиференціюємо п'яте
рівняння по змінній і віднімемо від нього восьме
рівняння останньої системи. Одержимо, що 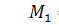 .
.
В силу того, що з шостого рівняння отримаємо, що 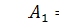
Будемо мати систему у
спрощеному вигляді:
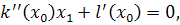
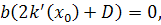
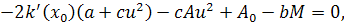
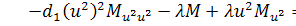 (2.2.2)
(2.2.2)
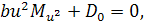

Перше рівняння отриманої
системи розщепимо по . Будемо мати, що 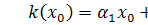 . Звідси бачимо, що
. Звідси бачимо, що 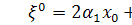 , а
, а 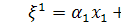 .
.
У другому рівнянні можливі 2
випадки:
. 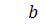
З шостого рівняння слідує, що
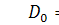 А отже
А отже 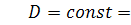
Розглянемо випадок, коли 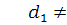
З п'ятого рівняння останньої
системи знайдемо  , проінтегрувавши його по
, проінтегрувавши його по  :
:
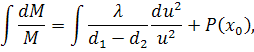
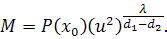
Взявши похідні по першого і другого порядку та
підставивши їх значення і значення в четверте рівняння, отримаємо
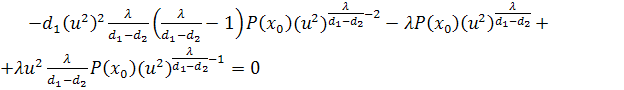
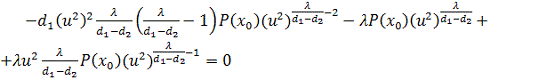 .
.
Так як 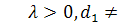 , функція
, функція 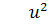 , то можна скоротити отримане
рівняння на
, то можна скоротити отримане
рівняння на 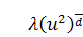 . Остаточно будемо мати
. Остаточно будемо мати
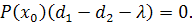
Можливі 2 випадки:
) 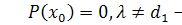
В цьому випадку =0.
Третє рівняння можна
розщепити по . Будемо мати
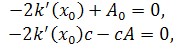
тобто
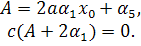
Підставимо значення з першого рівняння останньої
системи в друге:
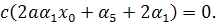
Розщепимо отримане рівняння
по:

Знову можливі 2 випадки:
а) 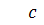
В цьому випадку 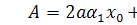 . Тоді
. Тоді
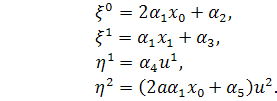
б) 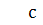
Тоді будемо мати наступну
систему
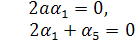
Можливі 2 випадки:
б1) 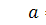
Тоді 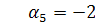 а
а 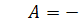 . Будемо мати
. Будемо мати
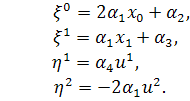
б2) 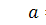
Тоді 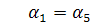 і
і  . Отримаємо
. Отримаємо
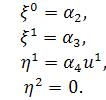
) 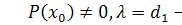
Підставимо значення 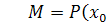 в останнє рівняння системи (2.2.2).
Будемо мати
в останнє рівняння системи (2.2.2).
Будемо мати
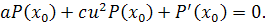
Розщепимо останнє рівняння
по:
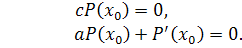
Так як 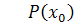 , то . Рівняння
, то . Рівняння 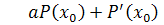 проінтегруємо по:
проінтегруємо по:
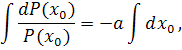
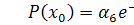 .
.
Тоді  .
.
Отримаємо
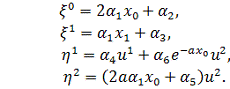
.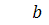
В цьому випадку  З шостого рівняння системи (2.2.2)
бачимо, що
З шостого рівняння системи (2.2.2)
бачимо, що 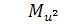 . Тоді з п'ятого рівняння випливає,
що =0.
. Тоді з п'ятого рівняння випливає,
що =0.
Третє рівняння можна
розщепити по . Будемо мати
тобто


Підставимо значення з першого рівняння останньої
системи в друге:
Розщепимо отримане рівняння
по :
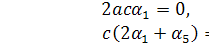
Можливі 2 випадки:
а)
В цьому випадку . Тоді
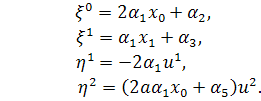
б)
Тоді будемо мати наступну
систему

Знову можливі 2 випадки:
б1)
Тоді а . Будемо мати
б2)
Тоді і . Отримаємо
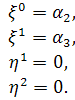
У випадку, коли можна ввести заміну  . Звідси
. Звідси 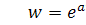 . Підставимо значення у друге рівняння системи (2.1.1).
Будемо мати
. Підставимо значення у друге рівняння системи (2.1.1).
Будемо мати

Доданок 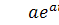 взаємознищується. Перепозначимо
взаємознищується. Перепозначимо 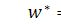 . Отримаємо
. Отримаємо
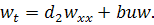
Тому в цьому випадку .
Остаточно отримаємо такі
випадки:
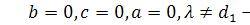
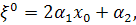



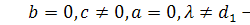


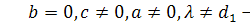

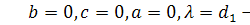
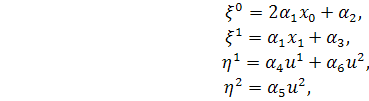
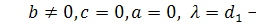
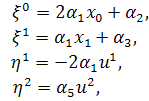

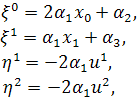
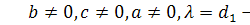
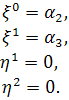
§2.3 Аналіз отриманих алгебр
інваріантності
Комутатором двох операторів i  називається оператор, який є
результатом дії
називається оператор, який є
результатом дії
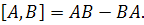
Для отриманих семи випадків
побудуємо базиси алгебр інваріантності і комутатори.
)
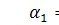
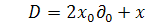
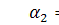
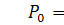
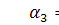
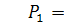
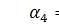
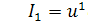
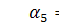
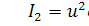
|
|
|
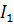
|
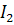
|
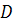
|
|
0 0 0 0 2
|
|
|
|
0
|
0
|
0
|
0
|
|
|
|
0
|
0
|
0
|
0
|
0
|
|
|
0
|
0
|
0
|
0
|
0
|
|
|
-2-000
|
|
|
|
|
2)
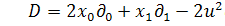
|
|
|
|
|
|
0 0 0 2
|
|
|
|
0
|
0
|
0
|
|
|
|
0
|
0
|
0
|
0
|
|
|
-2-00
|
|
|
|
3)
4)
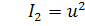 ,
,
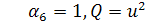 ,
,
|
|
|
|
|
Q
|
|
|
|
0
|
0
|
0
|
0
|
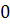
|
2
|
|
|
0
|
0
|
0
|
0
|
0
|
|
|
|
0
|
0
|
0
|
0
|
0
|
0
|
|
|
0
|
0
|
0
|
0
|
Q
|
0
|
|
Q
|
0
|
0
|
0
|
-Q
|
0
|
0
|
|
|
-2-0000
|
|
|
|
|
|
5)
,
|
|
|
|
|
|
0 0 0 2
|
|
|
|
0
|
0
|
0
|
|
|
|
0
|
0
|
0
|
0
|
|
|
-2-00
|
|
|
|
6)
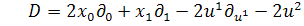
)
Розділ 3. Редукція системи
диференціальних рівнянь з частинними похідними до системи звичайних
диференціальних рівнянь
Розглянемо випадок, коли і проведемо редукцію по системі
рівнянь:
Перейдемо до змінних 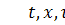 . Тоді
. Тоді
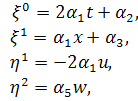
В нашому випадку система
(2.1.1) набере вигляду
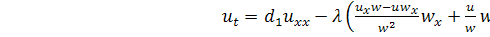
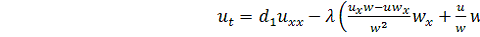 , (3.1.1)
, (3.1.1)
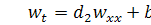 .
.
Складемо систему Ейлера -
Лагранжа :
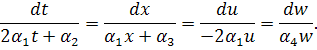
Розглянемо 2 випадки:
) 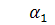
В цьому випадку система Ейлера -
Лагранжа буде мати вигляд:
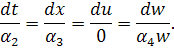
Проінтегруємо співвідношення 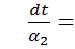 . Отримаємо
. Отримаємо
 .
.
називається інваріантною змінною.
Проінтегрувавши
співвідношення 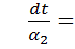 і
і 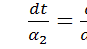 будемо мати
будемо мати

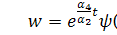
Отримаємо наступну систему

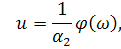
Дана система називається
анзацом, який редукує систему (3.1.1) до системи звичайних диференціальних
рівнянь. Візьмемо похідні від функцій і:
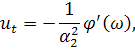
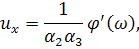
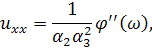
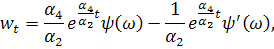
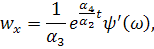
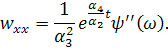
Підставимо знайдені похідні в
систему (3.1.1). Будемо вважати, що функції та залежать від:
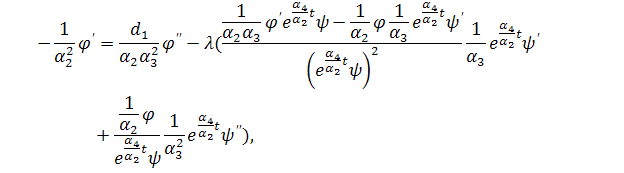
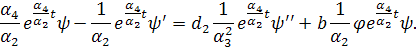
Скоротимо отриману систему на
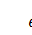 . Будемо мати наступну систему
. Будемо мати наступну систему
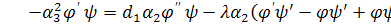 (3.1.2)
(3.1.2)
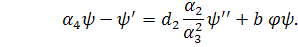
Система (3.1.2) є редукованою системою
з більшої розмірності в меншу.
2) 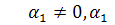
Cистема Ейлера - Лагранжа буде мати
вигляд:
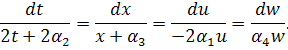
Проінтегрувавши
співвідношення 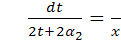 , знайдемо інваріантну змінну :
, знайдемо інваріантну змінну :
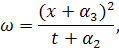
З співвідношення 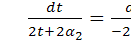 , скоротивши на
, скоротивши на  , знайдемо :
, знайдемо :
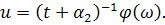
З співвідношення 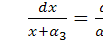 знайдемо
знайдемо  :
:
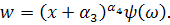
Отримаємо абзац у наступному
вигляді:
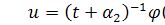
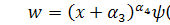
Візьмемо похідні від функцій і :
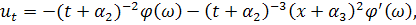
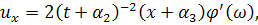
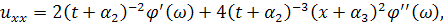
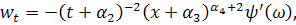
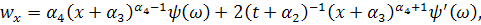
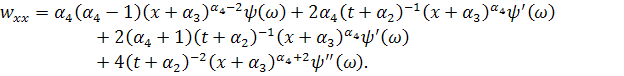
Підставивши знайдені похідні в
систему (3.1.1) і провівши аналогічні спрощення як у попередньому випадку,
отримаємо:

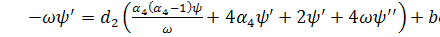 (3.1.3)
(3.1.3)
Зведемо систему (3.1.2) до
нормального виду. З другого рівняння цієї системи виразимо 
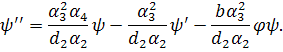
Значення підставимо в перше рівняння системи
(3.1.2) і знайдемо:
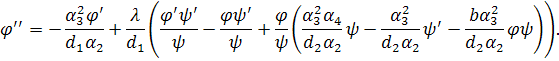
Проведемо наступну заміну 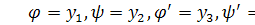 . Будемо мати систему
. Будемо мати систему

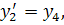
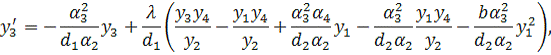
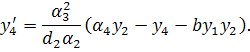
Остання система є системою
рівнянь (3.1.2) у нормальному вигляді. Дану систему можна досліджувати на
стійкість методами Ляпунова та Пуанкаре якісної теорії для звичайних
диференціальних рівнянь.
Аналогічно зводиться система
(3.1.3) до нормального вигляду.
Висновки
При написанні роботи можна зробити
висновок про те, що математичне моделювання відіграє важливу роль в біології.
Для побудови математичних моделей конкретних абіотичних і біотичних процесів,
що відбуваються в природних системах, необхідно також знати співвідношення, що
визначають потоки речовини і енергії як у систему так і з системи залежно від
стану окремих компонент цієї системи та навколишнього середовища.
У даній роботі було досліджено
математичну модель хемотаксису за допомогою методу Лі, який ґрунтується на
знаходженні та застосуванні операторів алгебр інваріантності (симетрій Лі)
розглядуваного нелінійного рівняння з частинними похідними для побудови його
точних розв'язків. У даній роботі запропоновано спеціальні анзаци, які
дозволяють провести редукцію системи ДРЧП, яка описує процес хемотаксису, до
системи ЗДР.
Робота складається з трьох розділів.
У першому розділі описане значення хемотаксису в математичній біології і в
життєвих процесах живих організмів та виведена загальна модель для опису
процесу хемотаксису. Другий розділ складається з трьох параграфів, в яких
показане виведення системи визначальних рівнянь, розв'язана отримана система
визначальних рівнянь, а також проаналізовані алгебри інваріантності, які були
одержані при розв'язанні СВР (обчислені комутатори і базиси алгебр
інваріантності). Третій розділ присвячений побудові анзаців максимальних алгебр
інваріантності математичної моделі хемотаксису, за допомогою яких проведена
редукція системи диференціальних рівнянь з частинними похідними до системи
звичайних диференціальних рівнянь.
Список використаної літератури
. Adler J. Chemotaxis in
Bacteria // Ann. Rev. Biochem. 1975. Vol. 44. P. 341 - 356.
2. Britton N. F. Essential
Mathematical Biology. - Berlin: Springer, 2003. - 335 P.
. Cherniha R. New
ansatze and exact solutions for nonlinear reaction-diffusion equations arising
in mathematical biology / Proceedings of the Second International Conference
"Symmetry in Nonlinear Mathematical Physics", Kyiv, 7-13 July 1997. -
Kyiv, In-te Math., 1997. - P. 138-146.
. Lucilla Corrias a ,
Benoit Perthameb . Asymptotic decay for the solutions of the parabolic -
parabolic Keller - Segel chemotaxis system in critical spaces. Mathematical and
Computer Modelling 47 (2008) 755 - 764.
. Murray J. D.
Mathematical Biology. - Berlin: Springer, 1989. - 767 P.
. Murray J. D.
Mathematical Biology, II. - Berlin: Springer, 2003. - 801 P.
. Дужин С. В.,
Чеботаревский Б. Д. От орнаментов до дифференциальных уравнений. - Минск :
Вышэйшая школа, 1988. - 254 с.
. Завальский Л. Ю.
Кинетический анализ хемотаксиса бактерий // Биофизика. 1988. Т. 33, №2. С. 328
- 332.
. Ибрагимов Н. Х. Азбука
группового анализа. // Новое в жизни, науке, технике. Сер. Математика. Кибернетика.
1989. - №8. - 48 с.
. Ибрагимов Н. Х. Группы
преобразований в математической физике. - М.: Наука, 1983. - 280 с.
. Компьютеры и
нелинейные явления. - М.: Наука, 1988. - 192 с.
. Лагно В.І., Спічак
С.В.,Стогній В.І. Симетрійний аналіз рівнянь еволюційного типу. Київ: Ін-т
математики НАН України, 2002. - 360 с.
. Овсянников Л.В.
Груповой анализ дифференциальных уравнений. М: Наука. - 1978. - 400 с.
. Скулачев В. П.
Электродвигатель бактерий // Соросовский Образовательный Журнал. 1998. №9. С. 2
- 7.
. Фущич В.И., Штелень
В.М., Серов Н.И. Симетрийный анализ и точные решения нелинейных уравнений
математической физики. Киев: Наук. думка. - 1989. - 336 с.
. Черніга Р.М.
застосування одного конструктивного методу для побудови неліївських розв'язків
нелінійних еволюційних рівнянь// Укр. мат. журн. - 1997. - Т.49, N 6, - С.
814-827.
хемотаксис анзац симетрія математичний
Додаток
Софус Лі
Лі Маріус Софус (17.XII.1842 -
18.II.1899) - норвезький математик. Народився в Ейд (неподалік Норфьордейде). В
1865 році закінчив університет в Христианіїї. За працю "Про представлення
уявних чисел в геометрії" отримав (1869 р.) стипендію для поїздки в
європейські наукові центри (Берлін, Париж).
В 1871 році за працю "Про один
клас геометричних перетворень" присуджена вчена ступінь доктора філософії.
У 1877 - 1886 рр. професура в
університеті Христианії. З 1886 р. - професор Лейпцигського університету.
Роботи присвячені теорії груп і
диференціальної геометрії. Автор класичної теорії неперервних груп (називається
теорією групи Лі), яка розвинулась згодом в загальну теорію неперервних груп.
Перші роботи (1871-72рр.) відносились суто до геометричних питань. Однак уже в
1872 р. Лі звернувся до теорії диференціальних рівнянь, ввів у неї поняття і
методи n-вимірної геометрії.
Теорія груп Лі, яка виникла, що
виникла з прагнення внести об'єднуючий початок і встановити загальні точки зору
в найрізноманітніших відділах математики, зробила глибокий вплив на подальший
розвиток теорії диференціальних рівнянь, алгебри, основ геометрії, топології і
теоретичної фізики. В результаті робіт Лі (і Ф. Клейна) геометрія була
перебудована на основі теоретико-групових перетворень.
Міжнародна премія ім.
М.І.Лобачевского присуджена в 1897 році за роботу по застосуванню теорії груп
для обґрунтування геометрії Лобачевского.
Анрі Пуанкаре
Анрі Пуанкаре народився 29 квітня
1854 року в містечку Сіте-Дюкаль поблизу Нансі (Лотарингія, Франція). Його
батько, Леон Пуанкаре, був професором медицини в Університеті Нансі.
Математична діяльність Пуанкаре
носила міждисциплінарний характер, завдяки чому за тридцять з невеликим років
своєї напруженої творчої діяльності він залишив фундаментальні праці практично
у всіх областях математики.
Роботи Пуанкаре, опубліковані
Паризькою Академією наук в 1916-1954, складають 10 томів. Це праці з топології,
теорії ймовірності, теорії диференціальних рівнянь, теорії автоморфних функцій,
неевклідової геометрії. Після захисту докторської дисертації, присвяченої
вивченню особливих точок системи диференціальних рівнянь, Пуанкаре написав ряд
мемуарів під загальною назвою «Про криві, визначені диференціальними
рівняннями». Ним уведені методи малого параметра, нерухомих точок, рівнянь у
варіаціях, розроблена теорія інтегральних інваріантів.
Пуанкаре належать багато важливих
для небесної механіки праць про стійкість руху і про фігури рівноваги
гравітуючої рідини, що обертається. Пуанкаре вперше ввів в розгляд автоморфні
функції і детально їх досліджував. Також він вперше ввів основні поняття
комбінаторної топології, такі як числа Бетті, фундаментальну групу, довів
формулу, що зв'язує число ребер, вершин і граней n-вимірного поліедра , дав
перше точне формулювання інтуїтивного поняття розмірності. В області
математичної фізики Пуанкаре досліджував коливання тривимірного континууму,
вивчив ряд задач теплопровідності, а також різні задачі в галузі теорії
потенціалів, електромагнітних коливань.
Ім'я Пуанкаре безпосередньо
пов'язане з успіхом теорії відносності. У 1905 році він написав твір «Про
динаміку електрона», в якому розвинув математичні наслідку «постулату
відносності».
- золота медаль королівського
астрономічного товариства .
- медаль Кетрін Брюс Тихоокеанського
Астрономічного Товариства.
Помер 17 липня 1912 року в Парижі.
Похований в сімейній гробовці на столичному кладовищі Монпарнас.