Макромицеты отделов Basidiomycota и Ascomycota защитных лесополос вдоль железной дороги Краснодар–Кореновск
Министерство
образования и науки Российской Федерации
Федеральное
государственное бюджетное образовательное учреждение
высшего
профессионального образования
КУБАНСКИЙ
ГОСУДАРСТВЕННЫЙ УНИВЕРСИТЕТ
Кафедра
биологии и экологии растений
ДИПЛОМНАЯ
РАБОТА
МАКРОМИЦЕТЫ
ОТДЕЛОВ BASIDIOMYCOTA И ASCOMYCOTA ЗАЩИТНЫХ ЛЕСОПОЛОС ВДОЛЬ ЖЕЛЕЗНОЙ ДОРОГИ
КРАСНОДАР-КОРЕНОВСК
Работу
выполнила О.В. Чернова
Факультет
биологический
Специальность
020201 Биология
Нормоконтролёр,
препод.
А.М.
Иваненко
Краснодар
2012
Реферат
Работа выполнена на 44 страницах машинописного
текста, состоит из введения, 4 глав, заключения и списка использованной
литературы объёмом в 44 литературных источников и двух приложений на 3 листах.
Работа проиллюстрирована 4 оригинальными цветными фотографиями в приложении.
Ключевые слова: МАКРОМИЦЕТЫ, ОТДЕЛ BASYDIOMYCOTA,
ОТДЕЛ ASCOMYCOTA,
ЛЕСОПОЛОСА, ЖЕЛЕЗНАЯ ДОРОГА, ФИТОЦЕНОЗ, АССОЦИАЦИЯ.
Целью данной работы является изучение
макромицетов отделов Basidiomycota
и Ascomycota защитных лесополос
вдоль железной дороги Краснодар - Кореновск.
Макромицеты отделов Basidiomycota
и Ascomycota защитных лесополос
вдоль железной дороги Краснодар - Кореновск, остаются мало изученными на
территории Динского района. Данные, приводимые в литературе противоречивы и малочисленны.
Мы впервые попробовали уточнить видовой состав, физико-географические условия
для обитания данных представителей, биоэкологические, экологические и
морфологические особенности, а так же на основе наших исследований провели
биоэкологический, таксономический, морфологический, анализы и определили
встречаемость и верность видов макромицетов отделов Basidiomycota
и Ascomycota в исследуемом
районе, провели фенологическое исследование.
Содержание
Введение
1. Обзор литературы
2. Физико-географическая
характеристика района исследований
1
Географическое
положение и рельеф Динского района
2
Климат
3
Почвы
4
Растительность
3. Материал и методы исследований
.1 Методы исследований
.2 Материал исследований
. Макромицеты отделов Basidiomycota
и Ascomycota защитных лесополос
вдоль железной дороги Краснодар - Кореновск
.1 Видовой состав и таксономический
анализ
.2 Биоэкологический анализ
.3 Морфологический анализ
.4 Встречаемость и верность видов
.5 Фенологические наблюдения
Заключение
Библиографический список
Приложение А
Введение
Грибы - особая форма жизни, царство живой
природы, объединяющее эукариотические организмы, сочетающие в себе некоторые
признаки, как растений, так и животных. Это одна из множества наибольших и
разнообразнейших групп живых организмов, появившаяся более миллиарда лет назад
и постепенно ставшая неотъемлемой частью всех водных и наземных экосистем.
Весьма велико биологическое и экологическое разнообразие грибов. В соответствии
с современными оценками, на Земле существует от 100 до 250 тысяч видов грибов,
но некоторые семейства грибов исследованы не достаточно. Они присутствуют во
всех экологических нишах - в воде, на суше. Играют важную роль в биосфере,
разлагая всевозможные органические материалы. Многие виды грибов активно
используются человеком в пищевых, медицинских и иных целях. Очень важна их роль
в природе как микоризообразователей. Однако следует отметить, что многие грибы
являются вредными для хозяйственной деятельности человека. Большой вред наносят
лесному хозяйству, поражая как растущие деревья, так и деловую древесину. Среди
грибов имеются паразиты человека и животных [Бурова, 1991].
Степень изученности видового состава той или
иной местности не однородна. Некоторые семейства царства грибов на немногих
территориях нашей станы изучены достаточно полно, другие слабее и их необходимо
рассматривать как для выяснения теоретического, так и практического значения
[Черновол, 2004]. К такой территории относится степная часть Северного Кавказа
микофлора, которой изучена недостаточно полно. Встречаются лишь отрывочные
сведения о макромицетах отделов Basidiomycota и Ascomycota защитных лесополос.
Недостаточно сведений по распространению и видовому составу царства грибов
макромицетов отделов Basidiomycota и Ascomycota защитных лесополос вдоль
железной дроги Краснодар - Кореновск.
Поэтому целью нашей работы стало исследование,
макромицетов отделов Basidiomycota
и Ascomycota защитных лесополос
вдоль железной дороги Краснодар - Кореновск. В задачи нашей работы входило:
1.Определить состав макромицетов отделов Basidiomycota
и Ascomycota защитных лесополос
вдоль железной дороги Краснодар - Кореновск и провести таксономический анализ;
2. Выявить
лимитирующие и топические факторы среды обитания макромицетов отделов Basidiomycota
и Ascomycota защитных лесополос
вдоль железной дороги Краснодар - Кореновск;
3. Выявить
биоэкологические и морфологические особенности макромицетов отделов Basidiomycota
и Ascomycota защитных лесополос
вдоль железной дороги Краснодар - Кореновск. И провести биоэкологический и
морфологический анализы;
4. Выявить
ядовитые группы макромицетов отделов Basidiomycota и Ascomycota защитных
лесополос вдоль железной дороги Краснодар - Кореновск;
5. Определить
следующие геоботанические характеристики, а именно: обилие по шкале Друде,
верность по шкале Браун-Бланке;
6. Провести
фенологические наблюдения и выявить сроки образования плодовых тел у видов
макромицетов изучаемых отделов Basidiomycota
и Ascomycota во флоре защитных
лесополос вдоль железной дороги Краснодар - Кореновск.
1. Обзор литературы
Грибы - обширная группа организмов,
насчитывающая около 100 тысяч видов. Данное царство встречается в самых
разнообразных местообитаниях: в воде, почве, на гниющих остатках животных и
растений. Грибы - неотъемлемый компонент всех процессов протекающих в
биоценозах. Велика так же их роль в круговороте веществ. Грибы наряду с
бактериями участвуют в почвообразовании, а в частности, в образовании
плодородного слоя почвы - гумуса [Аристамбекова,
2003].
Появились грибы у самих истоков органической
жизни в архейской эре около миллиарда лет назад. Произошли они от примитивных
одноклеточных жгутиковых и безжгутиковых амебовидных флагелят. Расцвет достигли
70-25 миллионов лен назад, в палеогеновый период. Указания на то, что грибов на
Земном шаре более 300 тысяч видов, не далеки от истины. На сегодняшний день
описано чуть более 65 тысяч грибов. Каждый год добавляется ещё тысяча видов, в
основном имеющих микроскопические размеры, то есть низших грибов микромицетов.
Высших же - макромицетов - на Земле около 15 тысяч видов, в Европе - 4,5
тысячи, в России - 3 тысячи [Черновол, 2004].
Наука, занимающаяся изучением грибов, получила
название - микология. Развитие микологии в России связано с именем выдающегося
ученого Михаила Степановича Воронина, которого с полным правом считают отцом
русской микологии. М. С. Воронин внес серьезный вклад в изучение циклов
развития ряда грибов, впервые обнаружил ловчие кольца на мицелии хищных грибов.
Учеником и продолжателем работ М. С. Воронина стал выдающийся миколог, исследователь
циклов развития многих грибов академик С. Г. Навашин [Жарбинский, 1987].
Для грибов лучше умеренно влажное и теплое, не
знойное лето. Вовсе не годится суховейное и дождливое лето, а если засушили
грибницу жаркие июнь и июль, тогда, лишь августовские и сентябрьские дожди
могут исправить дело: грибы пойдут под осень [Юдина, 1999].
По данным из некоторых источников не может
укрыться тот факт, что наибольшее число самых разных грибов растет в лесистой
местности и особенно в конце лета, и осенью. Однако видовой состав грибов
заметно различается в зависимости от характера местности [Двин, 2001].
Каждая древесная порода, так или иначе, связана
со «своими» микоризными грибами обычно присутствующими в лесах, где эта порода
господствует [Долетов, 2004].
В тополевых рощах чаще всего встречаются Amanita
albescens, Tricholoma
populinum, Boletus
chrysenteron [Янсен, 2004].
В дубовых рощах и зарослях белой акации, на
равнинах, холмах и в предгорьях чаще всего встречаются Amanita
phalloites, Amanita
muscaria, Amanita
rubescens [Васильева, 1939].
В смешанных лесах (дуб, граб, орешник) чаще
других встречаются Liccinum
carpini и Boletus
edulis [Юдин, 2003].
В более сухих дубовых рощах на не высоких горных
склонах часто попадаются съедобные цезарьский гриб (Amanita
cesanea), различные виды
сыроежек, рыжиков, волнушек, моховиков, боровиков [Янсен, 2007].
Очень богаты грибами каштановые леса с
вересковым и черничным пролеском. Здесь чаще
всего
встречаются
Amanita muscaria, Amanita rubescens, Russula emetica, Russula lepiola, Russula
cyanoxantra. Кроме того,
в
каштановых
лесах
можно
найти
грибы
семейства
болетовых
из
которых
наиболее
распространены:
Boletus edulus, Boletus pinicola, Suillus badius, Boletus subtomentosus и
Boletus lunidus [Зуев,
1988].
На коре
всех
вышеупомянутых
деревьев
живут
также
и
«сидячие»
грибы,
как
Fistulina hepatica и различные
грибы
из
семейства
полипоровых
- Grifola fronolosa, Lactiponus sulphuneus, Polyporus sguamosus - плодовые
тела
которых
съедобны.
Кроме
названных видов есть и Ramaria,
Clavaria, а также некоторые
аскомицеты: Pexixa и Helvella
[Черновол, 2004].
Особо нужно выделить трюфели. Это микоризные
подземные грибы, среди которых выделяются особо ценный белый трюфель - Tuber
magnatum, живущий в
симбиозе с дубом [Шемаханова, 1962].
При определении абиотических и топических
факторов для обитания грибов макромицетов отделов Basidiomycota
и Ascomycota защитных лесополос
вдоль железной дороги Краснодар - Кореновск, следует отметить то, что
своеобразная геологическая история, сложность физико-географических условий и
влияние человека с древнейших времён создали в этом крае разнообразный
растительный покров [Тильба, 1981].
Равнины северо-запада сейчас почти целиком
распаханы. Некогда это были степи, а в предгорьях - лесостепь. О характере естественной
растительности, господствовавшей здесь, можно судить теперь лишь по небольшим
её участкам, сохранившимся по оврагам, балкам и крутым берегам рек. Это были
ковыльные степи, луговые степи и варианты солончаковых степей [Тильба, 1981].
Многие авторы, дают разное описание наиболее
благоприятных для грибов мест обитания. М. В. Горленко, Л. М. Гарибова, М. В.
Бондарцева [1986] в своей книге говорят о том, что, изучая общие вопросы по
распространению грибов, легко прийти к выводу о том, что на них влияют в
основном те же факторы, которыми определяется, распределение. Распространение
живых организмов вообще. М. Кнооп [2000], говорит о том, что на живые
организмы, в том числе и на грибы влияют и такие пока еще недостаточно
изученные факторы, как земной магнетизм, интенсивность космической радиации,
атмосферное давление. С. Д. Додик [2001] утверждает, что одним из основных
факторов, определяющих расселение, распространение грибов, является наличие
подходящего субстрата: гумусовой почвы, лесной подстилки, древесины и различных
растительных остатков. Каждый из вышеперечисленных авторов выделяет главные для
себя аспекты, но в чем-то они все правы. Температура, влажность, интенсивность
осадков, освещенность, субстраты, за счет которых организмы питаются, состав атмосферы,
в которой грибы развиваются, и взаимодействие с другими организмами - важнейший
фактор, определяющий распространение данного царства - макромицетов.
В термальных водах и грязях можно найти
теплостойкие грибы, которые выдерживают температуру до 55°С, а в полярной
тундре встречаются грибы, переносящие зиму при минус 60°С. Для одних видов свет
может быть фактором, ускоряющим рост, у других (у навозника, например) при
свете рост замедляется [Федоров, 1994].
Что же касается местопроизрастания грибов, и их
топических пристрастий, то С. К. Понамарев [1987], указывает на то, что
грибница со склона не перейдет на террасу и уже никогда не покинет таковую. К.
Пименов [2006] утверждает что, отправляясь в незнакомый район, прежде всего,
следует отыскивать террасы или плоские водораздельные участки с разбросанными
по ним холмиками высотою 5-20 м. Почему-то именно такие формы рельефа особенно
полюбились грибам. По словам Ч. Д. Жарбинского [1987], на террасах грибы отдают
предпочтения вершинам холмов, а так же тыловым швам - это линия, по которой
терраса соединяется с нависающим над ней склоном и ригелям, бровкам над
уходящими вниз склонами.
Топические особенности грибов макромицетов
отделов Basidiomycota
и Ascomycota защитных лесополос
вдоль железной дороги Краснодар - Кореновск, было установлено, что сведений,
касающиеся данного вопроса мало.
Из некоторых источников [Федоров, 1994], можно
увидеть, что грибы отдают предпочтение кислым почвам. Д. П. Зуев [1960],
утверждает, что данные представители в большом количестве встречаются над
песчаниками и аргиллитами, и будут избегать известняков и мергелей. Хорошим
индикатором кислотности почв, как утверждает Г. И. Сержанина [1967], является
древесная растительность, например каштаны, любят кислые почвы. Причем, как
говорит Л. Н. Васильева [1939], следует учесть то, что на южной стороне
(склоне) будут произрастать дубы и грабы, а северной стороне (склону) отдают
предпочтение каштановые и буковые леса. Грибы, образующие микоризу только с
определенными одним или несколькими породами деревьев следует искать именно
вблизи этих деревьев.
Проще с ксиллофитами, как утверждает Ф. В.
Федоров [1990]: кого любят на том и селятся. Так, съедобная печеночница
встречается у основания стволов каштана и дуба, гериций иглистый на дубе. По
словам М. В. Горленко [1981], гастеромицеты - Дальдиния концентрическая,
вырастаю исключительно на валежнике из веток ясеня.
Из внешних абиотических условий очень важной
является температура, на это указывает А. В. Юдин [2003]. В статье Д. П.
Кассанелли, М. В. Нагалевского, С. Б. Криворотова, В. Я. Нагалевского [Редкие
виды грибов...,2003] утверждается, что большинство макромицетов отделов Basidiomycota
и Ascomycota теневыносливы и
влажнолюбивы (Hypholota,
Pholiota, Strapharia),
одноко, некоторые виды и рода светолюбивы (Psilocibe,
Stopharia). Интервал
температур, в которое вмещается все грибное царство, как утверждает М. В.
Горленко [1981], от 0 до 35°С. Но по Д. П. Зуеву [1960] наиболее благоприятны
для них температуры 20-30°С. По словам Ф. В. Федорова [1990], некоторые грибы,
предпочитают более низкие температуры, вот почему эти холодолюбивые грибы -
психрофиллы выбирают время парада ранней весной или поздней осенью, маслята,
например: ранней весной, в марте, и поздней осенью, в ноябре. По той же
причине, по словам В. Черновола [2004], в конце марта - начале апреля
появляются первые сморчки, сыроежки. И всю зиму при положительных температурах
хотя бы 1-5°С можно собирать на поваленных стволах бука вешенки-чинарики.
На пастбищах с хорошо дренированной песчаной легкой
почвой можно встретить много разных грибов, среди которых чаще всего
встречается аскомицеты и базидиомицеты из родов Lepista,
Lepiota, и Aganicus.
Виды рода Lepiota
чаще всего встречаются на почве, богатой гумусом, и на старых навозных кучах.
Здесь же часто можно найти грибы из рода Coprinus.
На известковых почвах чаще встречается Tricholoma
gambosum. Равнинные луга и
пастбища на склонах холмов - подходящие места для таких мелких грибов как Hygrocybe
lacta, Conica
coccinea [Сабатье, 2002].
He следует забывать
о таких родах грибов, как Psathynella,
Strophania, Lacnymania,
к которым относятся все сапротрофы, все они пластинчатые базидиомицеты. На
лугах и пастбищах иногда встречаются и грибы рода Nolanca
[Федоров, 1990].
По некоторым данным можно судить так же и о
биоэкологическом разнообразии макромицетов отделов Basidiomycota
и Ascomycota. Ф. Двин [2001],
отмечает, что подавляющее большинство грибов - это сапрофиты, питающиеся
органическим веществом отмерших организмов и растительными остатками, гниющими
корнями, разлагающейся лесной подстилкой, состоящей из ежегодно опадающих
отмерших частей древесно-кустарниковой растительности и отмирающей массы
почвенного покрова непосредственно влияет на запас и свойства подстилки, на
обогащение верхних горизонтов почвы минеральными веществами и на другие
процессы, упавшими на землю ветками и кусками коры, экскрементами травоядных
животных, реже - мертвыми насекомыми.
Другой не менее редко встречающейся
биоэкологической группой являются грибы паразиты [Додик, 2001]. Эти грибы живут
за счет живых растительных организмов, в частности, за счет взрослых деревьев:
они либо внедряются в случайные трещины, либо переносятся в виде спор
насекомыми, проедающими ходы в коре [Юдин, 2003]. Многим грибам для развития
необходимо, что бы их гифы соединялись с корешками растений. Это явление,
именуемое микоризным симбиозом, дает представление об экологии грибов
[Звонарев, 2008].
Отдел Basidiomycota
- это типичные грибы лесной местности. У них имеется более или менее развитая
ножка иногда просто цилиндрическая, иногда утолщенная или суживающаяся к
основанию, а бывает и изогнутая или плоская, уходящая в почву вместе с мицелием
разной длины [Двин, 2001]. Как утверждает Ф. В. Федоров [1994], поверхность
ножки может быть гладкая, морщинистая, комковатая, с прожилками, желобчатая. Е.
И. Коваленко, Н. Н. Коваленко, А. Е. Коваленко [1978], указывают на то, что
внутренность у ножки обычно плотная, но не редки случаи, когда она бывает полой
или частично полой. На ножке иногда имеется пленчатое кольцо или выпуклый
войлочный валик - все это следы специальных защитных оболочек, то есть общего
или частичного покрывала, обволакивающего молодое плодовое тело [Кнооп, 2000].
По мере роста последнего общее покрывало разрывается, и от него остаются следы
на ножке в виде вольвы, колец, поясков [Горленко, 1981]. Иногда вольва
оставляет свои следы в средней или верхней части ножки, приобретая форму
плотного кольца, нередко бесследно исчезающего [Зуев, 1988].
Говоря о морфологии отдела Basidiomycota
важно обратить внимание, по словам Ф. Двина [2001], на место и характер
соединения между ножкой и шляпкой. Оно может быть центральным, более или менее
сдвинутым от центра.
В процессе развития гриба форма шляпки очень
изменчива: вначале она может быть шарообразной, но по мере созревания
уплощается или даже ставится воронкообразной [Лебедева, 1949].
Как утверждает М. В. Горленко [1986],
поверхность шляпки может быть гладкой, волнистой, желобчатой, покрытой толстыми
или тонкими слоями слизи, потрескавшейся, блестящей, липкой, или сухой и
зернистой.
К данному отделу относится также такое семейство
как Amanitacea
(мухаморовые), или аманитовые. К ним относятся многочисленные виды, в основном,
микоризных грибов с широкой шляпкой, к которой иногда прирастают белёсые
колечки, оставшиеся от общего покрывала. Ножка цилиндрическая, высокая, внизу с
утолщением [Черновол, 2004].
Не менее обширное семейство, относящееся к
отделу Basidiomycota,
является семейство Russulaceae
(сыроежковые). В зрелом виде у данных грибов шляпка в центре бывает вогнутой
или даже воронкообразной. Поверхность шляпки гладкая, иногда липкая. Это
семейство включает многочисленные виды сапрофитов и микоризных грибов,
принадлежащих к родам Russula
и Lactanius [Черновол, 2004].
У семейства лепиоптовых шляпка может быть разных
размеров покрытая толстым слоем чешуек. Больбитиевые (Bolbitiaceae)
имеют вытянуто-коническую с гладкой, блестящей поверхностью. Ножка обычно
тонкая, выпуклая [Андрест, 1984].
Что же касается отдела Ascomycota,
то, как утверждает Ф. Двин [2001], заметный переход от ножки к шляпке
встречается очень редко, во всяком случае, у них никогда не бывает характерной
для отдела Basidiomycota
«классической» формы зонтика. У некоторых представителей данного класса, как
указывает Д. П. Зуев [1960], иногда можно различить ложную ножку и ложную
шляпку (моршеловые, гельвеловые).
Широко известно семейство Morchellaceae
(сморчковые) относится к отделу. Это семейство имеет четко разделенную шляпку и
ножку. Шляпка имеет более или менее коническую форму, ячеистая, полая [Кнооп,
2000].
Другое семейство Rezizaceae
(пецицевые) имеет представители, у которых относительно небольшие плодовые тела
и лишь у немногих видов есть ножка в основании [Зуев, 1988].
Лишенные хлорофилла и любого другого типа
фотосинтетического пигмента, грибы зависят от источника органического питания
уже готового к потреблению. Их способность к развитию прямо или косвенно
связанна с хлебными растительными остатками живых организмов, выделениями
животных [Гарибова, 1981].
Что же касается вопроса о группе ядовитых грибов
макромицетов отдела Basidiomycota
и Ascomycota, а именно, сколько
грибов насчитывается на территории Северного Кавказа, то на этот вопрос имеется
достаточно литературных источников. Поэтому можно так же судить о
распространении ядовитых макромицетов отделов Basidiomycota
и Ascomycota защитных лесополос
вдоль железной дороги Краснодар - Кореновск [Кнооп, 2000].
На территории Европы встречается коло 80 видов
грибов, употребление которых в пищу всегда или при определенных условиях может
вызвать неприятное явление или серьезное нарушения функций организма вплоть до
опасных для жизни [Горленко, 1986]. По данным М. Н. Сергеевой [2000], на
Северном Кавказе насчитывается около двух тысяч ядовитых грибов. Эти грибы М.
В. Горленко, Л. М. Гарибова, М. В, Бондарцева [1981], поделили на три большие
группы: не съедобные, условно съедобные и ядовитые. М. В. Вишневский [2004],
указывает на то, что к не съедобным грибам относятся те, которые не содержат
ядовитых веществ, но имеют неприятный запах или чаще горький или едкий вкус.
Как утверждают многие авторы, например М. В.
Вишневский [2001], эти грибы не вызывают отравлений, но могут быть причиной
неприятных ощущений или легких нарушений пищеварения. С. Г. Мусселиус [2007],
говорит о том, что наиболее распространенным и часто по ошибке собираемым
грибом является горчак или желтый гриб. П. Хардинг [2002] считает, что наиболее
распространенным не съедобным грибом, в сухих хвойных, реже в лиственных лесах,
является перечный гриб, близкий родственник маслят. По книге Е. И. Коваленко,
Н. Н. Коваленко, А. Е. Коваленко [1978], наиболее распространенным грибом
Северного Кавказа, произрастающего на живых стволах и валежнике лиственных
деревьев, является чешуйчатка обыкновенная. В книге К. Л. Даддингтона [1959],
самыми распространенными не съедобными макромицетами являются некоторые
млечники (серо-розовый, шиповатый), сыроежки (пятнистая, рвотная).
Следует упомянуть и следующую группу грибов -
это условно съедобные грибы. В работе Ф. В. Федорова [1994], ей уделяется большое
внимание. По его словам в данную группу относят тех представителей, которые
содержат ядовитые или сильно раздражающие вещества, которые, однако,
разрушаются или удаляются при соответствующей обработке гриба. Как утверждает
Ч. Д. Жарбинский [1987], условно съедобные грибы появляются в лесах Северного
Кавказа ранней весной - это сморчки и строчки. К условно съедобным грибам, по
словам Л. Н. Васильевой [1939], принадлежат так же многочисленные млечники и
некоторые сыроежки, имеющие жгучий или горький вкус (скрипица, перечный груздь,
волнушки) или неприятный запах (валуй).
В группу ядовитых грибов С. Г. Мусселиус [2007],
объединяет грибы, в плодовых телах которых на всех стадиях развития содержатся
ядовитые вещества - токсины, вызывающие отравление. В Европе к наиболее опасным
относят 20-25 видов грибов, среди них смертельно ядовитые - бледная поганка,
мухомор вонючий и весенний, некоторые волокинницы. Практически все эти виды
встречаются в лесах и лесостепях Северного Кавказа. Следует отметить то, что
неотъемлемым элементом при дальнейшем изучении макромицетов отделов Basidiomycota
и Ascomycota защитных лесополос
вдоль железной дороги Краснодар - Кореновск, станет ландшафт.
Степная растительность некогда покрывала всю
северную равнинную часть края, относящуюся к степной зоне. Более 70% площади
степей распахано, занято сельскохозяйственными культурами [Тильба, 1981].
Следует так же, отметить, что большинство лесополос являются деятельностью рук
человека. Поэтому следует так же обратить внимание на происходящие изменения
связанные с микроклиматическими условиями, выражающиеся в понижении
освещённости, температуры воздуха и почвы. С повышением сомкнутости насаждений
происходят изменения и в гидрологическом режиме насаждений [Хардинг, 2002]. Это
всё может привести к необратимым изменениям, как во флоре, так и в царстве
грибов.
Исторически сложилось так, что в России, как ни
в одной стране грибы стали традиционным национальным блюдом, причем сушеные и
соленые издавна были предметом экспорта во многие страны. Любят у нас грибы и
сейчас, причем ряды грибников все больше и больше пополняются городским
населением. Теперь уже стали обычными у нас специальные поезда в пригородные
грибные места, выезды в лес по грибы на автобусах целыми коллективами. К
сожалению, еще можно порой видеть глубокую колею прямо на лесной подстилке -
колыбели грибов, а легковой автомобиль под деревом на поляне, где быть ему не
положено [Федоров, 1990].
В ходе исследования нами были использованы
различные методы: биоэкологический и морфологический анализ, метод прямого
учета, глазомерный метод, метод пробных площадок и описательный метод. В ходе
исследований изучались морфологические особенности встречаемых макромицетов
отделов Basidiomycota
и Ascomycota (размер ножки,
шляпка, окраска) [Ефремов, 2001].
Для выявления фитоценотической роли макромицетов
отделов Basidiomycota
и Ascomycota лесополос вдоль
железной дороги Краснодар - Кореновск, мы пользовались общепринятой
геоботанической методикой определения обилия и верности.
Представители отделов Basidiomycota
и Ascomycota являются
компонентами различных фитоценозов. Следовательно, в жизни леса и в общем
круговороте веществ и в природе эти грибы играют немаловажную роль.
Популяции многих макромицетов отделов Basidiomycota
и Ascomycota предпочитают
лиственные леса, но не редкость их встретить так, же и в хвойных лесах. Так в
буково-разнотравных ассоциациациях часто встречаются Amanita
phalloides, Amanita
junguillea, Russula
lepiola. В дубовых рощах и
в зарослях белой акации чаще всего встречаются Amanita
phalloidess, Amanita
muscaria, Amanita
nubescens, Lactarius
torminosus. В тополевых рощах
с подлеском гниющей травы, встречаются Amanita
nubescens, Lactarius
suboluleis, Tricholoma
populium, Boletus
chrysentenon. В более сухих
дубовых рощах может попасться Amanita
cesanea, различные виды
сыроежек, рыжиков, моховиков, боревиков [Смирнов, 1979].
Каждый день, проведенный в лесу, нужно
записывать: в каком лесу был, какие грибы в нем встретил, когда, какие явления
сопутствовали (цвели, плодоносили травы, кусты, деревья), много ли было грибов.
Такие наблюдения называют фенологическими (от двух греческих слов: phaino -
являю и logos - понятие, наука) [Янсен, 2004].
На основе таких записей можно составить для лета
данного года календарь явлений в лесу: первое появление грибов того или иного
вида, время их массового урожая, когда встречались в лесу последние грибы,
каков был урожай грибов. Если такие наблюдения велись ряд лет, можно составить
средний многолетний календарь для данного района. Такой календарь будет иметь
не только познавательное, но и практическое значение. По нему можно заранее
составлять план сбора первых грибов, определять время массового урожая грибов
для заготовок их в данной местности [Мусселиус, 2007].
2. Физико-географическая
характеристика района исследований
.1 Географическое положение и рельеф
Динского района
макромицет basidiomycota лесополоса
биоэкологический
Динской район расположен к северо-востоку от г.
Краснодара в центральной части Краснодарского края. Станица Динская - районный
центр, находится в 30 км от г. Краснодара, в 4 км от трассы Республиканского
значения Краснодар - Ростов-на-Дону. Район занимает удобное географическое
положение, граничит с Усть-Лабинским, Кореновским, Тимашевским, Калининским
районами, городом Краснодаром, Республикой Адыгея.
Территорию Динского района пересекают три
железнодорожные магистрали общегосударственного значения. В станице Динской
работает два крупных железнодорожных узла. Кроме того, район имеет широко
развитое автодорожное сообщение. По его территории проходит автомобильная
дорога федерального значения «Дон» и две дороги субъекта федерации: «Краснодар
- Ейск» и «Темрюк - Краснодар - Кропоткин» [Ситникова, 2000].
Динской район расположен в степной зоне края,
поэтому не имеет лесов естественного происхождения. Лесонасаждения представлены
полезащитными и водораздельными лесными полосами [Дубинин, Чекрыгина, 1988].
Степная растительность некогда покрывала всю
северную часть края, относящуюся к степной зоне и составляющую около половины
её территории [Тильба, 1981].
2.2 Климат
Динской район находится в пограничной полосе
двух климатов: континентального и средиземноморского. И поэтому ему присущи
короткие теплые зимы и жаркое лето с достаточным количеством дождей.
Существенное влияние на климат оказывает близость Черного и Азовского морей
[Ситникова, 2000].
При восточных и северо-восточных ветрах летом
устанавливается сухая и жаркая погода. Зимой эти же ветры приносят холод. Ветры
юго-западных направлений летом приносят прохладу и дожди, зимой - осадки в виде
снега и дождя [Дубинин, Чекрыгина, 1988].
Влияют на климат также и местные ветры: фены,
бризы [Ситникова, 2000].
2.3 Почвы
Наибольшую площадь в районе занимают черноземные
почвы, они относятся к так называемым западно-предкавказским черноземом, отличаются
плодородием. Мощность почвенного слоя достигает 1,5-2 м. По шкале оценки
биологической продуктивности земли, Динской район имеет повышенный
биоклиматический потенциал [Ситникова, 2000].
Основная часть площади черноземов занята
сельскохозяйственными культурами. Климатические условия являются важнейшим
фактором биологической продуктивности сельскохозяйственных растений. Наиболее
существенным в том плане являются: тепло, свет и влага. Она позволяют при малом
количестве гумуса достигать высоких урожаев зерновых колосовых культур
[Дубинин, Чекрыгина, 1988].
Минерально-сырьевые ресурсы представлены глиной,
песком и подземными пресными водами [Дубинин, Чекрыгина, 1988].
2.4 Растительность
Естественная растительность представлена
лугово-степными видами. Сейчас в местах, где когда-то произрастали степные
ковыли, пырей, вика, тимофеевка и прочее степное разнотравье, на распаханных
землях колосятся хлеба [Ситникова, 2000].
По берегам рек среди степного разнотравья
произрастают мелкие поросли древесной растительности (орешник, дикий миндаль),
имеются так же непроходимые заросли колючего терна. На водоразделах равнин
можно встретить дуб, дикую грушу, яблоню, татарский клен, бузину, терн,
шиповник, ежевику. По долинам рек - вербу, кустарниковую иву, черный и белый тополь,
ольху. На песчаных берегах растет солодка, синеголовник приморский, люцерна,
тимофеевка. Кое-где можно встретить редкие заросли деревьев и кустарников,
среди которых встречаются дуб, дикая груша, лещина, шиповник, держидерево,
ежевика. В связи с достаточным увлажнением, реки богаты ряской, водорослями.
Берега рек поросли зарослями камыша, рогоза [Ситникова, 2000].
Имеются так же лесополосы искусственного
насаждения, в которых произрастает древесная растительность - клен, ясень,
тополь белый, яблоня, груша, ежевика, бузина, терн [Дубинин, Чекрыгина, 1988].
3. Материал и методы исследований
.1 Методы исследований
Основные методы, которые были использованы нами
при изучении макромицетов отделов Basidiomycota и Ascomycota защитных лесополос
вдоль железной дороги Краснодар - Кореновск, проходящих по станице Динской -
это биоэкологические и морфологические анализы, метод прямого учета,
глазомерный метод, метод пробных площадок, описательный метод, методы
проведения анализов почвенного раствора. Во время наших исследований был собран
материал, который мы проанализировали методом прямого учета [Воронов, 1973].
Для приближенного учета обилия макромицетов отделов Basidiomycota и Ascomycota
защитных лесополос вдоль железной дороги Краснодар - Кореновск, был использован
глазомерный метод абсолютного учета и шкала Друде:− грибы доминируют;
СОР3 − грибы очень обильны;
СОР2 − грибы обильны;
СОР1 − грибы довольно обильны;−
грибы редкие;− грибы единичны;− единичный экземпляр.
По пятибалльной шкале Браун − Бланке изучили
показатель степени приуроченности видов (верность видов) к определенным
ассоциациям [Воронов, 1973]:
− виды, случайно попавшие в эту ассоциацию
и чуждые ей;
− виды, встречающиеся во многих
ассоциациях;
− виды, встречающиеся во многих
ассоциациях, но все же предпочитающие данную;
− виды, встречающиеся преимущественно в
данной ассоциации;
− виды, встречающиеся исключительно в
данной ассоциации.
По В. Я. Нагалевскому [1987], применяли метод
пробных площадок, в тех случаях, когда ассоциации занимаю значительную площадь.
Конкретные участки ассоциации были описанны на так называемой пробной площадке
определенных размеров. Нами будут изучаться макромицеты отделов Basidiomycota и
Ascomycota защитных лесополос вдоль железной дороги Краснодар - Кореновск. Вот систематический
список данных классов:
Отдел: Ascomycota
Класс: Pyrenomycetes
Порядок: Clavicipitales
Порядок: Hypocreales
Класс: Discomycetes
Порядок: Pezizales
Порядок: Tuberales
Отдел: Basidimycota
Класс: Holobasidiomycetes
Порядок: Hymenomycetidae
Порядок: Aphyllophorales
Пopядoк:Agaeicales
Подкласс: Gasteromycetidae
Порядок: Melanogasterales
Порядок: Hysterangiales
Порядок: Sclerodermatales
Порядок: Lycoperdales
Порядок: Nidulariales
Класс: Heterabasidiomycetes
Порядок: Auriculariales
Порядок: Tulasnellales
Порядок: Tremellales
Порядак: Dacrymycetales
Данная классификация была взята из книги Н. П.
Черепановой [2005].
3.2 Материал исследований
В нашей работе объектами исследования являются
макромицетов отделов Basidiomycota
и Ascomycota защитных лесополос
вдоль железной дороги Краснодар - Кореновск. В данных лесополосах нами было
найдено 7 семейств отдела Basidiomycota
и 2 семейства отдела Ascomycota.
Далее приводится список видов встречающихся макромицетов отделов Basidiomycota
и Ascomycota:
Отдел Basidiomycota:
Семейство: Amanitacea
(мухоморовые)
Amanita
phalloides (бледная поганка)
Amanita
verna (весенняя поганка)
Amanita
virosa (мухомор вонючий)
Amanita
excelsea (мухомор высокий)
Amanita
ovoidea (мухомор
яйцевидный)
Amanita
muscaria (мухомор красный)
Amanita
rubescens (мухомор розовый)
Amanita
spissa (мухомор толстый)
Семейство: Tricholomataceae
(рядовковые)
Tricholoma
sulphereum (рядовка
сернистая)
Tricholoma
populinum (рядовка
тополевая)
Семейство: Russulaceae
(сыроежковые)
Russula
aeruginia (сыроежка зеленая)
Russula
emetica (сыроежка рвотная)
Russula
declorans (Сыроежка
сереющая)
Russula
pectinata (сыроежка
гребенчатая)
Russula
vesca (сыроежка пищевая)
Семейство: Paxilaceae
(свинушковые)
Ompholotius
olearius (свинушка
маслянистая)
Семейство: Aganicaceae
(шампиньоновые)
Aganicus
augustus (шампиньон
августовский)
Семейство: Boletaceae
(болетовые)
Boletus
luridus (дубовик
обыкновенный)
Boletus
purpureus (моховик
пурпурный)
Boletus
subtomentosus (моховик зеленый)
Boletus
chrysenteron (моховик пестрый)
Семейство: Coprinaceae
(навозниковые)
Coprinus
atramentarius (навозник серый)
Coprinus
comatus (навозник белый)
Coprinus
disseminates (навозник
рассеянный)
Отдел Ascomycota:
Семейство: Pezizaceae
(пецицивые)
Otodea
onotica (ослиные уши)
Pezia
badia (пецица
коричневая)
Семейство: Sarcosomataceae
(саркосомевые)
Sarcosoma
cocсinea
(саркосома ярко-красная)
Исследования проводились в период с 2010 по 2011
год в Динском районе (ст. Пластуновская, ст. Платнировская) и городе Кореновск,
Краснодарского края.
Местом исследования стали защитные лесополосы
вдоль железной дороги Краснодар - Кореновск.
При установлении сезона образования плодовых тел
изучаемых нами макромицетов отделов Basidiomycota
и Ascomycota, использовался
материал фенологических наблюдений, проводимых в период 2010 - 2012 гг.
4. Макромицеты отделов Basidiomycota
и Ascomycota
защитных лесополос вдоль железной дороги Краснодар - Кореновск
.1 Видовой состав и таксономический
анализ
Как уже описывалось нами ранее, информации по
видовому составу представителей макромицетов отделов Basidiomycota
и Ascomycota защитных лесополос
вдоль железной дороги Краснодар - Кореновск, очень мало.
За время проведения исследований нами было
выявлено 7 семейств грибов отдела Basidiomycota,
включающие в себя 24 вида и 2 семейства грибов отдела Ascomycota,
включающие 3 вида, на территории, показанной на рисунке А 1.
Самыми многовидовыми семействами, встретившимися
на территории наших исследований, стали семейства отдела Basidiomycota:
Amanitaceae, Russulaceae,
Bolbitiaceae и семейство отдела
Ascomycota: Pezizaceae.
4.2 Биоэкологический анализ
В ходе исследований нами было выделено 7
экологических групп, которые встречаются среди изученных нами семейств.
Наиболее часто встречаемой экологической группой являются сапрофиты и почвенные
сапрофиты. К ним относится наибольшее количество видов выше описанных нами
семейств (Tricholoma sulphereum, Amanita phalloides, Russula emeticа, Russula
pectinata, Aganicus augustus, Boletus purpureus, Coprinus comatus). Не редкость
встретить представителей относящихся к капрофиллам, селящихся на гниющих
растительных остатках и на навозе (Coprinus
atramentarius, Coprinus
disseminates). Некоторые
представители могут относиться как к капрофиллам так и к ксиллосапрофитам (Coprinus
atramentarius). Имеются
представители и психрофиллов, предпочитающие низкие температуры (Amanita
verna). Не редкость
встретить так же представителей относящихся к ксиллофитам, грибов селящихся у
основания различных древесных пород (Tricholoma
populinum, Ompholotius
olearius, Otodea
onotica, Pezia
badia, Sarcosoma
cocсine).
Семейство Amanitacea
мухаморовые, или аманитовые включает в себя многочисленные виды, в основном,
микоризных грибов (Amanita
excelsea, Amanita
ovoidea, Amanita
spissa).
4.3 Морфологический анализ
Наши наблюдения показывают, что морфологические
особенности видов грибов отделов Basidiomycota
и Ascomycota касаются в
основном формы шляпки, наличия или отсутствия ножки и ее формы.
Отдел Basidiomycota:
Семейство Amanitacea
(мухоморовые) в основном имеет сферическую (Amanita
phalloides) или
полусферическую (Amanita
verna) шляпку. Ножка
длинная, тонкая (Amanita
excelsа) иногда
цилиндрическая (Amanita
rubescens). Amanita
ovoidea имеет плоскую
опушенную нитевидными отростками шляпку и крепкую, покрытую пушком ножку.
Другой представитель (Amanita
muscaria) имеет плоскую с
углублением в центре шляпку, цилиндрическую ножку. Amanita
spissa имеет плоскую
шляпку и утолщающуюся книзу ножку. Семейство Tricholomataceae
(рядовковые) имеет распростертую шляпку, тонкую ножку (Tricholoma
populinum). Некоторые
представители имеют распростертую и слегка приплюснутую шляпку, ножка
булавовидной формы (Tricholoma
sulphereum). Семейство Russulaceae
(сыроежковые) имеет плоскую шляпку, гладкую толстую ножку (Russula
aeruginia), у некоторых
представителей шляпка вдавленная и бугристая, ножка плотная, крепкая (Russula
emeticа). Russula
declorans имеет
полусферическую шляпку, мясистую, плотную ножку. Russula
pectinata имеет слегка
вдавленную в центре шляпку, толстую, но очень ломкую ножку. Что же касается Russula
vesca то данный
представитель имеет шляпку с углублением в центре и волнистые края, ножка
цилиндрическая. Семейство Paxilaceae
(свинушковые) имеет плотную воронкообразную шляпку и высокую, заостренную у
основания ножку (Ompholotius
olearius). Семейство Aganicaceae
(шампиньоновые) имеет мясистую иногда с бугорком в центре шляпку, ножка
крепкая, цилиндрическая (Aganicus
augustus). Семейство Boletaceae
(болетовые) имеет распрастертую шляпку, крепкую, клубневидную ножку (Boletus
luridus), некоторые
представители имеют выпуклую со слегка волнистыми краями шляпку и толстую,
булавовидную ножку (Boletus
purpureus). Иногда можно
встретить представителей с выпуклой, с дольчатыми краями шляпкой и толстой
цилиндрической ножкой (Boletus
subtomentosus), не редкость
встретить представителя с плокой шляпкой и удлиненной, цилиндрической ножкой (Boletus
chrysenteron). Семейство Coprinaceae
(навозниковые) имеет колокольчатую шляпку и цилиндрическую утолщенную книзу
ножку (Coprinus
comatus). Coprinus
comatus имеет
узкоколокольчатую шляпку и довольно высокую ножку. Другой представитель (Coprinus
disseminates) имеет
колокольчатую шляпку и длинную, тонкую ножку.
Отдел Ascomycota:
Семейство Pezizaceae
(пецицивые) имеет завернутые края плодового тела, ножка не большая (Otodea
onotica). Другой
представитель имеет плодовое тело в форме чаши и рудиментированную ножку (Pezia
badia). Семейство Sarcosomataceae
(саркосомевые) имеет плодовое тело в форме чашечки и очень короткую ножку (Sarcosoma
coceinea).
4.4 Встречаемость и верность видов
Для оценки встречаемости (обилию) и
приуроченности видов относящихся к наиболее часто встречающимся макромицетам
отделов Basidiomycota
и Ascomycota в исследуемой нами
местности, мы на маршрутных ходах глазомерным методом по усовершенствованной
трехбалльной шкале Друде [Воронов, 1973] определяли встречаемость, (обилие)
видов макромицетов относящихся к отделам Basidiomycota
и Ascomycota. Одновременно
отвечали по шкале Браун-Бланке приуроченность (верность), наиболее часто встречающихся
видов макромицетов отделов Basidiomycota
и Ascomycota.
В таблице 1 отражены обобщенные материалы
исследований по ассоциациям макромицетов отдела Basidiomycota.
Таблица 1 - Обилие (по шкале Друде) и верность
(по шкале Браун - Бланке) видов отдела Basidiomycota
в растительных сообществах защитных лесополос вдоль железной дороги Краснодар -
Кореновск
|
Вид
|
Ассоциация
|
Обилие
|
Верность
|
|
Amanita phalloides (бледная
поганка)
|
дубово-бузиновая
|
COP1
|
3
|
|
дубово-разнотравная
|
SOL
|
2
|
|
ясенево-бузиновая
|
SP
|
1
|
|
A. verna (весенняя
поганка)
|
дубово-мертвоопадная
|
COP3
|
3
|
|
кленово-разнотравная
|
SP
|
2
|
|
кленово-злаково-разнотравная
|
SOL
|
2
|
|
A. virosa (мухомор
вонючий)
|
ясенево-разнотравна
|
COP1
|
2
|
|
глидичево-разнотравная
|
SOL
|
1
|
|
A. excelsea (мухомор
высокий)
|
робинево-овсяницевая
|
COP1
|
2
|
|
робинево-разнотравная
|
SOL
|
3
|
|
ясенево-мертвоопадная
|
SOL
|
2
|
|
А.
ovoidea
(мухомор
яйцевидный)
|
дубово-бузиновая
|
COP1
|
1
|
|
дубово-разнотравная
|
SOL
|
3
|
|
ясенево-бузиновая
|
COP3
|
3
|
|
A. muscaria (мухомор
красный)
|
кленово-разнотравная
|
COP1
|
2
|
|
ясенево-бузиновая
|
SOL
|
3
|
|
ясенево-разнотравна
|
SOL
|
2
|
|
А.
rubescens (мухомор
розовый)
|
дубово-разнотравная
|
SP
|
1
|
|
дубово-бузиновая
|
COP1
|
3
|
|
глидичево-разнотравная
|
SOL
|
3
|
|
Вид
|
Ассоциация
|
Обилие
|
Верность
|
|
Amanita spissa (мухомор
толстый)
|
робинево-разнотравная
|
SP
|
2
|
|
ясенево-разнотравна
|
SOL
|
2
|
|
кленово-разнотравная
|
COP1
|
1
|
|
Tricholoma sulphereum (рядовка
сернистая)
|
дубово-бузиновая
|
SOL
|
1
|
|
дубово-разнотравная
|
COP3
|
2
|
|
ясенево-бузиновая
|
SOL
|
1
|
|
T. populinum (рядовка
тополевая)
|
дубово-мертвоопадная
|
COP1
|
2
|
|
кленово-разнотравная
|
SOL
|
2
|
|
кленово-злаково-разнотравная
|
SP
|
3
|
|
Russula aeruginia (сыроежка
зеленая)
|
ясенево-разнотравна
|
SOL
|
2
|
|
глидичево-разнотравная
|
COP1
|
2
|
|
робинево-овсяницевая
|
SOL
|
1
|
|
R. emetica (сыроежка
рвотная)
|
робинево-разнотравная
|
COP1
|
1
|
|
ясенево-мертвоопадная
|
SOL
|
2
|
|
дубово-бузиновая
|
SP
|
1
|
|
R. declorans (сыроежка
сереющая)
|
дубово-разнотравная
|
SOL
|
3
|
|
ясенево-бузиновая
|
SOL
|
2
|
|
дубово-мертвоопадная
|
COP1
|
2
|
|
R. pectinata (сыроежка
гребенчатая)
|
COP3
|
1
|
|
кленово-злаково-разнотравная
|
SOL
|
3
|
|
ясенево-разнотравна
|
SP
|
3
|
|
R. vesca (сыроежка
пищевая)
|
глидичево-разнотравная
|
COP1
|
3
|
|
робинево-овсяницевая
|
SOL
|
3
|
|
дубово-бузиновая
|
SOL
|
2
|
|
Ompholotius
olearius (свинушка
маслянистая)
|
дубово-разнотравная
|
COP1
|
2
|
|
ясенево-бузиновая
|
SP
|
1
|
|
дубово-мертвоопадная
|
SOL
|
2
|
|
Вид
|
Ассоциация
|
Обилие
|
Верность
|
|
Aganicus augustus
(шампиньон августовский)
|
кленово-разнотравная
|
COP1
|
5
|
|
дубово-бузиновая
|
SOL
|
1
|
|
глидичево-разнотравная
|
SP
|
2
|
|
Boletus luridus (дубовик
обыкновенный)
|
глидичево-разнотравная
|
COP1
|
3
|
|
робинево-овсяницевая
|
SOL
|
1
|
|
ясенево-бузиновая
|
SP
|
2
|
|
B. purpureus (моховик
пурпурный)
|
дубово-мертвоопадная
|
COP3
|
2
|
|
кленово-разнотравная
|
SOL
|
2
|
|
кленово-злаково-разнотравная
|
COP1
|
2
|
|
B. subtomentosus (моховик
зеленый)
|
глидичево-разнотравная
|
COP1
|
1
|
|
робинево-овсяницевая
|
SOL
|
2
|
|
робинево-разнотравная
|
SP
|
2
|
|
B. chrysenteron (моховик
пестрый)
|
дубово-мертвоопадная
|
SOL
|
3
|
|
ясенево-бузиновая
|
SP
|
3
|
|
дубово-бузиновая
|
COP1
|
3
|
|
Coprinus atramentarius (навозник
серый)
|
дубово-разнотравная
|
SOL
|
2
|
|
ясенево-бузиновая
|
SP
|
2
|
|
глидичево-разнотравная
|
SOL
|
2
|
|
C. comatus (навозник
белый)
|
дубово-бузиновая
|
COP1
|
1
|
|
ясенево-бузиновая
|
SP
|
3
|
|
глидичево-разнотравная
|
SOL
|
1
|
|
C. disseminates (навозник
рассеянный)
|
глидичево-разнотравная
|
SOL
|
2
|
|
дубово-бузиновая
|
COP1
|
2
|
|
ясенево-мертвоопадная
|
COP3
|
3
|
Из данных приведенных в таблице 1 можно увидеть,
что популяция вида Amanita
phalloides довольно обильна в
дубово-бузиновой ассоциации (СОР1), в дубово-разнотравной грибы
единичны (SOL), в
ясенево-мертвоопадной грибы редки (SP).
Анализ приуроченности к определенным ассоциациям данного вида показал, что
данный вид предпочитает дубово-бузиновую ассоциацию, чуждой для данного вида
является ясенево-бузиновая ассоциация и довольно часто данный вид можно
встретить в дубово-разнотравной ассоциации.
Популяция вида Amanita
verna обильна в
дубово-мертвоопадной ассоциации (СОР3), в кленово-разнотравной грибы
редки (SP), единичны
в кленово-злаково-разнотравной ассоциации (SOL).
Анализ приуроченности к определенным ассоциациям данного вида показал, что
данный вид предпочитает дубово-мертвоопадную ассоциацию, довольно часто данный
вид можно встретить в кленово-злаково-разнотравной и кленово-разнотравной.
Популяция вида Amanita
virosa довольно обильна в
ясенево-разнотравной ассоциации (СОР1), в глидичево-разнотравной
ассоциации грибы единичны (SOL).
Анализ приуроченности к определенным ассоциациям данного вида показал, что
данный вид чуждый для глидичево-разнотравной ассоциации, наиболее часто его
можно встретить в ясенево-разнотравной ассоциации.
Популяция вида Amanita
excelsea довольно обильна в
робинево-овсяницевой ассоциации (СОР1), в робинево-разнотравной
ассоциации и ясенево-мертвоопадной грибы данного представителя встречаются
единично (SOL). Анализ
приуроченности к определенным ассоциациям данного вида показал, что данный вид
предпочитает робинево-разнотравную ассоциацию, наиболее часто его можно
встретить в ясенево-мертвоопадной и робинево-овсяницевой ассоциациях.
Популяция вида Amanita
ovoidea очень обильна в
ясенево-бузиновой ассоциации (СОР), в дубово-бузиновой грибы довольно обильны
(СОР1), в дубово-разнотравной данный вид встречается единично (SOL).
Анализ приуроченности к определенным ассоциациям данного вида показал, что
данный вид предпочитает дубово-разнотравную и ясенево-бузиновую ассоциации,
чуждой для данного вида является дубово-бузиновая ассоциация.
Популяция вида Amanita
muscaria довольно обильна в
кленово-разнотравной ассоциации (СОР1), в ясенево-бузиновой и
ясенево-разнотравной ассоциациях грибы единичны (SOL).
Анализ приуроченности к определенным ассоциациям данного вида показал, что
данный вид предпочитает ясенево-бузиновую ассоциацию, довольно часто данный вид
можно встретить в кленово-разнотравной и ясенево-разнотравной ассоциациях.
Популяция вида Amanita
rubescens довольно обильна в
дубово-бузиновой ассоциации (СОР1), в глидичево-разнотравной
представители данного вида единичны (SOL),
в дубово-разнотравной редки (SP).
Анализ приуроченности к определенным ассоциациям данного вида показал, что
данный вид предпочитает дубово-бузиновую и глидичево-разнотравную ассоциации,
чуждой для данного вида является дубово-разнотравная ассоциация.
Популяция вида Amanita
spissa довольно обильна в
кленово-разнотравной ассоциации (СОР1), в ясенево-разнотравной вид
встречается редко (SOL),
в робинево-разнотравной грибы редки (SP).
Анализ приуроченности к определенным ассоциациям данного вида показал, что
данный вид часто можно встретить в робинево-разнотравной и ясенево-разнотравной
ассоциациях, кленово-разнотравной ассоциация - чуждая для данного
представителя.
Популяция вида Tricholoma
sulphereum обильна в
дубово-разнотравной ассоциации (СОР), в дубово-бузиновой и ясенево-бузиновой
ассоциациях грибы единичны (SOL).
Анализ приуроченности к определенным ассоциациям данного вида показал, что
данный вид часто можно встретить в дубово-разнотравной ассоциации, в
дубово-бузиновая и ясенево-бузиновая ассоциации - чуждые.
Популяция вида Tricholoma
populinum довольно обильна в
дубово-мертвоопадной ассоциации (СОР1), в кленово-разнотравной грибы
единичны (SOL),
кленово-злаково-разнотравной ассоциации грибы данного вида редки (SP).
Анализ приуроченности к определенным ассоциациям данного вида показал, что
данный вид часто можно встретить в дубово-мертвоопадной и кленово-разнотравной
ассоциациях, предпочитает данный вид кленово-злаково-разнотравную ассоциацию.
Популяция вида Russula
aeruginia довольно обильна в
глидичево-разнотравной ассоциации (СОР1), в ясенево-разнотравной и
робинево-овсяницевой ассоциациях грибы единичны (SOL).
Анализ приуроченности к определенным ассоциациям данного вида показал, что в
ясенево-разнотравной и робинево-овсяницевой ассоциациях данный вид можно
встретить довольно часто, робинево-овсяницевая ассоциация - чуждая.
Популяция вида Russula
emetica довольна обильна в
робинево-разнотравной ассоциации (СОР1), в ясенево-мертвоопадной
грибы единичны (SOL),
в дубово-бузиновой редки (SP).
Анализ приуроченности к определенным ассоциациям данного вида показал, что
часто данный вид можно встретить в ясенево-мертвоопадной ассоциации, для
робинево-разнотравной и дубово-бузиновой ассоциаций виды чужды.
Популяция вида Russula
declorans довольно обильна в
дубово-мертвоопадной ассоцивции (СОР1), в ясенево-бузиновой и
дубово-разнотравной ассоциациях представители единичны (SOL).
Анализ приуроченности к определенным ассоциациям данного вида показал, что
часто данный вид можно встретить в ясенево-бузиновой и дубово-мертвоопадной
ассоциациях, дубово-разнотравную ассоциацию данный вид предпочитает.
Популяция вида Russula
pectinata очень обильна в
кленово-разнотравной ассоциации (СОР3), в кленово-злаково-разнотравной
грибы единичны (SOL),
в ясенево-разнотравной редки (SP).
Анализ приуроченности к определенным ассоциациям данного вида показал, что
данный вид предпочитает кленово-злаково-разнотравную и ясенево-разнотравную
ассоциации, кленово-разнотравная - чуждая.
Популяция вида Russula
vesca довольно обильна в
глидичево-разнотравной ассоциации (СОР1), в робинево-овсяницевой и
дубово-бузиновой грибы единичны (SOL).
Анализ приуроченности к определенным ассоциациям данного вида показал, что
данный вид предпочитает робинево-овсяницевую и глидичево-разнотравной
ассоциации, в дубово-бузиновой представители довольно часты.
Популяция вида Ompholotius
olearius довольно обильна в
дубово-мертвоопадной ассоциации (СОР), в кленово-разнотравной представители единичны
(SOL), в
кленово-злаково-разнотравной грибы редки (SP).
Анализ приуроченности к определенным ассоциациям данного вида показал, что
данный вид предпочитает кленово-злаково-разнотравную ассоциацию, в
кленово-разнотравнаой и дубово-мертвоопадной ассоциациях представителей данного
вида можно встретить довольно часто.
Популяция вида Aganicus
augustus довольно обильна в
кленово-разнотравной ассоциации (СОР1), в дубово-бузиновой
представители единичны (SOL),
в глидичево-разнотравной редки (SP).
Анализ приуроченности к определенным ассоциациям данного вида показал, что
данный вид предпочитает исключительно кленово-разнотравную ассоциацию, довольно
часто данный вид можно встретить в глидичево-разнотравной ассоциации, чуждой
является дубово-бузиновая ассоциация.
Популяция вида Boletus
luridus довольно обильна в
глидичево-разнотравной и ясенево-бузиновой ассоциации (СОР1), в
робинево-овсяницевой грибы единичны (SOL).
Анализ приуроченности к определенным ассоциациям данного вида показал, что
данный вид предпочитает глидичево-разнотравную ассоциацию, в ясенево-бузиновой
представителей можно встретить часто, робинево-овсяницевая - чуждая.
Популяция вида Boletus
purpureus очень обильна в
дубово-мертвоопадной ассоциации (СОР3), в
кленово-злаково-разнотравной представители довольно обильны (СОР1),
в кленово-разнотравной грибы единичны (SOL).
Анализ приуроченности к определенным ассоциациям данного вида показал, что
данный вид часто встречается как в дубово-мертвоопадной ассоциации,
кленово-злаково-разнотравной так и в кленово-разнотравной ассоциациях.
Популяция вида Boletus
subtomentosus довольно обильны
(СОР1) в глидичево-разнотравной ассоциации, в робинево-овсяницевой
грибы единичны (SOL),
робинево-разнотравной редки (SP).
Анализ приуроченности к определенным ассоциациям данного вида показал, что
данный вид часто можно встретить в робинево-овсяницевой и робинево-разнотравной
ассоциации, в глидичево-разнотравная - чуждая.
Популяция вида Boletus
chrysenteron довольно обильна в
дубово-бузиновой ассоциации (СОР1), в дубово-мертвоопадной
представители единичны (SOL),
в ясенево-бузиновой представители редки (SP).
Анализ приуроченности к определенным ассоциациям данного вида показал, что
данный вид предпочитает все три ассоциации.
Популяция вида Coprinus
atramentarius единична в
дубово-разнотравной и глидичево-разнотравной ассоциации (SOL),
в ясенево-бузиновой редки (SP).
Анализ приуроченности к определенным ассоциациям данного вида показал, что
данный вид можно встретить во всех трех ассоциациях. Популяция вида Coprinus
comatus довольно обильны
(СОР1) в дубово-бузиновой ассоциации, единичны в
глидичево-разнотравной ассоциации (SOL),
в ясенево-бузиновой ассоциации редки (SP).
Анализ приуроченности к определенным ассоциациям данного вида показал, что
данный вид предпочитает ясенево-бузиновую ассоциацию, чуждыми являются -
глидичево-разнотравная и дубово-бузиновая ассоциации.
Популяция вида Coprinus
disseminates очень обильна в
ясенево-мертвоопадной ассоциации (СОР), в дубово-бузиновой обильны (СОР), в
глидичево-разнотравной ассоциации редки (SP).
Анализ приуроченности к определенным ассоциациям данного вида показал, что
данный вид можно встретить часто в глидичево-разнотравной и дубово-бузиновой,
предпочитает данный вид ясенево-мертвоопадную ассоциацию.
В таблице 2 отражены обобщенные материалы
исследований по ассоциациям макромицетов отдела Ascomycota.
Таблица 2 - Обилие (по шкале Друде) и верность
(по шкале Браун - Бланке) видов отдела Ascomycota
в растительных сообществах защитных лесополос вдоль железной дороги Краснодар -
Кореновск
|
Вид
|
Ассоциация
|
Обилие
|
Верность
|
|
Otodea onotica (ослиные
уши)
|
глидичево-разнотравная
|
SOL
|
2
|
|
дубово-бузиновая
|
COP1
|
2
|
|
ясенево-мертвоопадная
|
SP
|
2
|
|
Peziza badia (пецица
коричневая)
|
глидичево-разнотравная
|
COP3
|
3
|
|
робинево-овсяницевая
|
SOL
|
2
|
|
робинево-разнотравная
|
SP
|
2
|
|
Sarcosoma cocсinea
(саркосома ярко-красная)
|
дубово-разнотравная
|
COP3
|
3
|
|
ясенево-бузиновая
|
SP
|
2
|
|
дубово-мертвоопадная
|
SP
|
2
|
Из данных приведенных в таблице 2 можно увидеть,
что популяция вида Otodea
onotica довольно обильна в
дубово-бузиновой ассоциации (СОР), в глидичево-разнотравной представители
единичны (SOL), в
ясенево-мертвоопадной редки (SP).
Анализ приуроченности к определенным ассоциациям данного вида показал, что
данный вид можно встретить часто в трех данных ассоциациях.
Популяция вида Peziza
badia очень обильна в
глидичево-разнотравной ассоциации (СОР), в робинево-овсяницевой представители
единичны (SOL), в
робинево-разнотравной редки (SP).
Анализ приуроченности к определенным ассоциациям данного вида показал, что данный
вид можно встретить часто в робинево-овсяницевой, робинево-разнотравной
ассоциациях, представители данного вида предпочитают глидичево-разнотравную
ассоциацию.
Популяция вида Sarcosoma
coccinea очень обильна в
дубово-разнотравной ассоциации (СОР), редки представители данного вида в
ясенево-бузиновой и дубово-мертвоопадной ассоциации. Анализ приуроченности к
определенным ассоциациям данного вида показал, что данный вид предпочитает
дубово-разнотравную ассоциацию, довольно часто данные представители встречаются
в ясенево-бузиновой и дубово-мертвоопадной ассоциации.
4.5 Фенологические наблюдения
Фенологические наблюдения проводили в период с
марта по октябрь с 2010 по 2012 гг. на территории Краснодарского края и
непосредственно в районе исследования - в защитных лесополосах вдоль железной
дороги Краснодар - Кореновск. Наблюдения велись в фазе образования плодовых
тел.
Установили сезон плодоношения видов изучаемых
нами отделов. Но этот показатель может очень сильно варьировать из года в год,
потому что напрямую зависит от различных экологических факторов, например
температуры, влажности.
По годам изменения в образовании плодовых тел у
разных представителей семейств макромицетов отдела Basidiomycota были, но в
среднем данные отражены на рисунке 1.
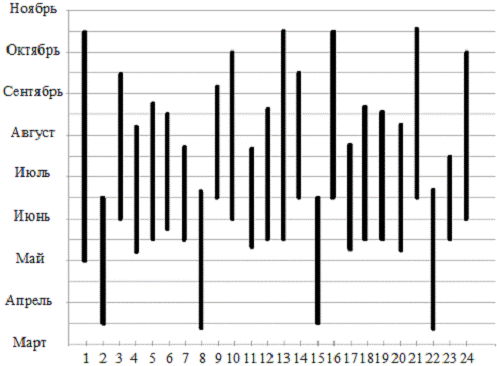
Рисунок 1 - Сезон образования плодовых тел
макромицетов отдела Basidiomycota защитных лесополос вдоль железной дороги
Краснодар - Кореновск.
- Amanita
phalloides (бледная поганка);
2 - Amanita
verna (весенняя
поганка); 3 - Amanita
virosa (мухомор вонючий);
4 - Amanita
excelsea (мухомор высокий);
5 - Amanita
ovoidea (мухомор
яйцевидный); 6 - Amanita
muscaria (мухомор красный);
7 - Amanita
rubescens (мухомор розовый);
8 - Amanita
spissa (мухомор толстый);
9 - Tricholoma
sulphereum (рядовка
сернистая); 10 - Tricholoma
populinum (рядовка
тополевая); 11 - Russula
aeruginia (Сыроежка
зеленая); 12 - Russula
emetica (сыроежка
рвотная); 13 - Russula
declorans (сыроежка
сереющая); 14 - Russula
pectinata (сыроежка
гребенчатая); 15 - Russula
vesca (сыроежка
пищевая); 16 - Ompholotius
olearius (свинушка
маслянистая); 17 - Aganicus
augustus (шампиньон
августовский); 18 - Boletus
luridus (дубовик
обыкновенный); 19 - Boletus
purpureus (моховик
пурпурный); 20 - Boletus
subtomentosus (моховик зеленый);
21 - Boletus
chrysenteron (моховик пестрый);
22 - Coprinus
atramentarius (навозник серый);
23 - Coprinus
comatus (навозник белый);
24 - Coprinus
disseminates (навозник
рассеянный).
Таким образом, видно, что время образования
плодовых тел у представителей семейств макромицетов отдела Basidiomycota
индивидуально, но массовое появление плодовых тел наблюдается в период с 3
декады мая по сентябрь.
К ранним срокам появления плодовых тел семейства
Amanitacea (мухоморовые),
исходя из данных рисунка 1, относятся Amanita
verna и Amanita
spissa. Вероятнее всего
это связанно с особенностями произрастания видов в весенний период когда почва
прогревается до температуры, необходимой для образования плодовых тел.
Сравнивая временные промежутки начала образования плодовых тел, можно сказать,
что у Amanita
verna, Amanita
spissa, Russula
vesca из семейства Russulaсеае
(сыроежковые) и Coprinus.
аtramentarius относящийся к
семейству Coprinасеае
(навозниковые), они совпадают. Самый длинный период плодоношения имеют Amanita
phalloides и Amanita
virosa из семейства Amanitacea
(мухоморовые), Tricholoma
populinum из семейства Tricholomataceae
(рядовковые), Russula
declorans из семейства Russulaceae
(сыроежковые) и Coprinus
disseminates из семейства Coprinaceae
(навозниковые). А Amanita
excelsea, Amanita
ovoidea, Amanita
muscaria, Amanita
rubescens из семейства Amanitacea
(мухоморовые), Russula
aeruginia, Russula
emetic из семейства Russulaсеае
(сыроежковые), Aganicus
augustus из семейства Aganicасеае
(шампиньоновые), Boletus
luridus, Boletus
purpureus, Boletus
subtomentosus из семейства Boletасеае
(болетовые) и Coprinus
comatus из семейства Сoprinасеае
(навозниковые), имеют самые короткие сроки плодоношения.
Массовое образование плодовых тел,
представителей семейств макромицетов отдела Basidiomycota, наблюдается в
наиболее благоприятный период - июнь, июль, август.
Что же касается представителей отдела
Ascomycota, то следует отметить, что по годам изменения наблюдались, но в
среднем данные отражены на рисунке 2.
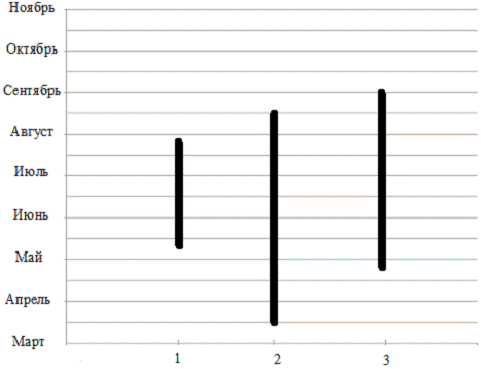
Рисунок 2 - Сезон образования плодовых тел
макромицетов отдела Ascomycota защитных лесополос вдоль железной дороги
Краснодар - Кореновск.
- Otodea
onotica (ослиные уши); 2 -
Peziza badia
(пецица коричневая); 3 - Sarcosoma
cocсinea
(саркосома ярко-красная).
Таким образом, видно, что время образования
плодовых тел у разных представителей семейств макромицетов отдела Ascomycota
индивидуально, но массовое появление плодовых тел наблюдается в период с 3
декады мая по сентябрь.
К ранним срокам появления плодовых тел относится
Peziza badia
из семейства Pezizaсеае
(пецицевые). Самый длинный промежуток плодоношения имеют Peziza
badia и Sarcosoma
cocсinea
из семейства Sarcosomataceae
(саркосомовые). А Otodea
onotica из семейства Pezizaсеае
(пецицевые) имеет самый короткий срок вегетации. Массовое образование плодовых
тел, представителей семейств макромицетов отдела Ascomycota, наблюдается в
наиболее благоприятный период - июнь, июль, август.
Заключение
По результатам работы сделаны следующие выводы:
1. Выявленный состав макромицетов относится к 2
отделам 9 семейств 17 родам и включает 27 видов.
. Лимитирующими факторами для исследуемых видов
являются: низкое увлажнение, сильное затенение и высокая температура.
Фитоценотически не благоприятны для развития макромицетов являются:
робинево-разнотравная и глидичево-разнотравная ассоциации.
. В процессе исследований выявлены 7
экологических групп макромицетов (сапрофиты, почвенные сапрофиты, капрофиллы,
ксиллосапрофиты, психрофиллы, ксиллофиты, микоризообразователи). Среди
исследованных видов морфологически преобладают шляпочные грибы с центральной
ножкой.
. Группа ядовитых макромицетов насчитывает 3
вида (Amanita
phalloides, Russula
emetic, Sarcosoma
coccinea).
5. Анализ встречаемости выявил значительное
участие следующих популяций: Coprinus
disseminates, Boletus
purpureus, Russula
pectinata, Tricholoma
sulphereum, Amanita
verna, Peziza
badia, Sarcosoma
coccinea. Анализ верности
видов выявил, что наиболее верным является распространение Aganicus
augustus в
кленово-разнотравной ассоциации. Большинство видов получили бал 3 и бал 1
получили те виды, которые случайно попали в чуждую им ассоциацию.
. Анализ фенологических наблюдений показал, что
наиболее ранние сроки плодоношения соответствуют 5 видам (Amanita
verna, Amanita
spissa, Russula
vesca, Coprinus
atramentarius, Peziza
badia), поздние сроки
плодоношения соответствуют 5 видам (Amanita
phalloides, Tricholoma
populinum, Russula
declorans, Ompholotius
olearius, Boletus
chrysenteron).
Библиографический список
1. Андрест Б. В.
Грибное лукошко. М., 1984. 230 с.
2. Аристамбекова Н. Г. Грибы.
М., 2003. 260 с.
. Бурова Л. Г. Загадочный мир
грибов. М., 1991. 196 с.
. Васильева Л. Н. Грибы
Кавказского заповедника. Казань, 1939. 116 с.
. Вишневский М. В. За грибами
с ноября по май. М., 2004. 267 с.
. Вишневский М. В.
Несъедобные, ядовитые и галлюциногенные грибы. М., 2001. 192 с.
. Воронов А. Г. Геоботаника:
учеб. пособие для ун-тов и пед. ин-тов. М., 1973. 384 с.
. Гарибова Л. В. В царстве
грибов. М., 1981. 230 с.
. Горленко М. В., Гарибова Л.
М., Бондарцева М. В. Всё о грибах. М., 1981. 280 с.
. Горленко М. В. Грибы СССР.
М., 1986. 303 с.
. Ефремов А. П. Смертоносные
растения и грибы. М., 2001. 290 с.
. Даддингтон К. Л. Хищные
грибы - друзья человека. М., 1959. 190 с.
. Двин Ф. Грибы. М., 2001.
304 с.
. Додик С. Д. Грибы
российских лесов. М., 2001. 320 с.
. Долетов Ю. К. секреты
заядлого грибника. М., 2004 с.
. Дубинин В. И., Черыгина
Т.А. Динская. Краснодар, 1988. 48 с.
. Жарбинский Ч. Д. По грибы.
М., 1987. 230 с.
. Звонарев Н. М. Грибы.
Собираем, выращиваем, заготовляем. М., 2008. 192 с.
. Зуев Д. П. Дары русского
леса. М., 1960. 190 с.
. Зуев Д. П. Дары русского
леса. М., 1988. 192 с.
. Кнооп М. Всё о грибах. М.,
2000. 254 с.
. Коваленко Е. И., Коваленко
Н. Н., Коваленко А. Е. Съедобные и ядовитые грибы Кубани. Краснодар, 1978. 175
с.
. Лебедева Л. А. Определитель
шляпочных грибов. М., 1949. 548 с.
. Мусселиус С. Г. Грибы
ядовитые. М., 2007. 254 с.
. Нагалевский В. Я. Учебная
практика по систематике растений с основами геоботанике: метод. указания.
Краснодар, 1987. 23 с.
. Нагалевский В. Я.,
Криворотов С. Б., Кассанелли Д. П. Редкие виды грибов Краснодарского края //
Актуальные вопросы экологии и охраны природы экосистем южных регионов России и
сопридельных регионов: материалы VII межресп. науч.-практ. конф. Краснодар,
2003 С. 20 - 25.
. Пименов К. Грибы в лесу, на
столе, в аптечке. М., 2006. 196 с.
. Пономарев С. К. Тебе
грибник. Липецк, 1987. 61 с.
. Сабатье Р. Грибы. М., 2002.
136 с.
. Сергеева М. Н. Грибы. М.,
2000. 263 с.
31. Ситникова Н. В.
Моя Кубань. Ростов н/Д, 2000. 208 с.
32. Смирнов В. И. Грибы. М.,
1979. 192 с.
. Съедобные и ядовитые грибы:
определитель / под ред. Г. И. Сержаниной. Минск, 1967. 183 с.
. Тильба А. П. Растительность
Краснодарского края: учеб. пособие. Краснодар, 1981. 84 с.
35. Федоров Ф. В.
Грибы: в 6 т. Т. 3. М., 1990. 360 с.
36. Федоров Ф. В. Грибы: в 6 т.
Т. 5. М., 1994. 364 с.
. Хардинг П. Грибы. М., 2002.
254 с.
38. Черновол В.
Грибное очарование лесов Кубани. Краснодар, 2004. 192 с.
39. Черепанова Н. П. Систематика
грибов: учеб. пособие. СПб., 2005. 344 с.
. Шемаханова Н. М. Микотрофия
древесных пород. М., 1962. 256 с.
. Юдин А. В. Большой
определитель грибов. М., 2003. 245 с.
. Юдина И. А. Советы
грибнику. М., 1999. 240 с.
. Янсен П. Все о грибах.
СПб., 2004. 265 с.
. Янсен П. Грибы. СПб., 2007.
192 с.
Приложение А
Карта Динского района
Рисунок А.1 - Схема Динского района