Модель индукции синтеза IgE
Модель индукции
синтеза IgE
Основная биологическая роль IgE
связана с его уникальной способностью связываться с поверхностью тучных клеток
и базофилов человека. Только эти клетки несут высокоаффинный гликопротеиновый
рецептор (относительная молекулярная масса =130 000 дальтон) для Fc-участка
молекулы IgE (FceRl).
На поверхности одного базофила имеются 40 000-100 000 рецепторов, которые
связывают от 5 000 до 40 000 молекул IgE
[3, 7]. На других клетках (В-лимфоцитах, макрофагах, эозинофилах и тромбоцитах)
были выявлены низкоаффинные рецепторы для IgE
(FceRII). Надмембранная
часть этих рецепторов легко поддается протеолитическому отщеплению от клеточной
мембраны, превращаясь в растворимый FceRII,
идентичный молекуле CD23,
обладающий способностью связывать IgE.
В связи с этой способностью та же молекула получила название «IgE
связывающий фактор>> (IgE-BF).
Этот фактор ингибирует связывание IgE
не только с FceRII, но и с FceRI,
участвуя, таким образом, в регуляции IgE-зависимых
аллергических реакций. Кроме того, этот фактор участвует в регуляции продукции IgE:
Т-хелперы, несущие FceRII-peцепторы
(CD23), продуцируют
фактор, усиливающий гликозилирование IgE-BF,
а Т-лимфоциты со свойствами супрессоров продуцируют фактор, ингибирующий
гликозилирование, гликозилированный IgE-BF
потенцирует, а негликозилированный фактор угнетает продукцию IgE.
Однако регуляция продукции IgE
- это гораздо более сложный процесс (Рисунок 1). Выявлена ассоциация
специфического IgE-ответа
с определенным HLA-DR
гаплотипом.
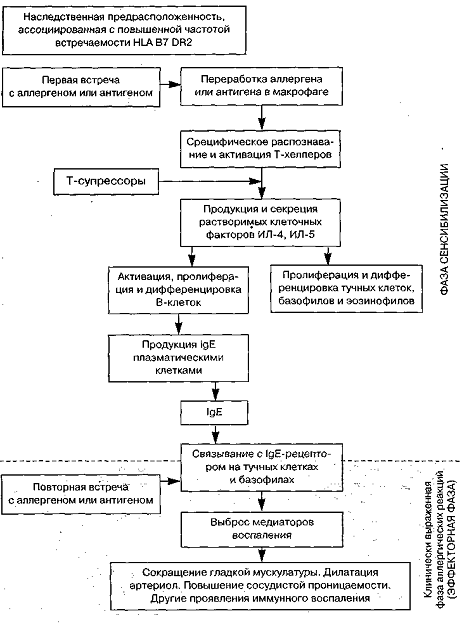
Рисунок 1. Схема продукции IgE,
её регуляции и IgE-опосредованных
эффектов.
Начальной фазой
антиген(аллерген)индуцированного этапа является активация В-лимфоцитов, которая
требует двух сигналов: от непосредственно распознающих: антигены Ig-рецепторов
В-лимфоцитов и от Т-хелперов. непосредственно и через цитокины, продуцируемые
активированными Т-хелперами. Последние являются наиболее важным звеном в
регуляции синтеза иммуноглобулинов, так как запускают также этапы пролиферации
и дифференцировки активированных В-лимфоцитов в плазматические
(антите-лопродуцирующие) клетки. На этом этапе синтез IgE.
по сравнению с иммуноглобулинами других классов, имеет некоторые особенности
(Рисунок 2). Среди цитокинов, контролирующих продукцию IgE,
есть два цитокина, оказывающих разнонаправленное действие на его синтез:
интерлей-кин-4 (IL-4)
стимулирует, а интерферон-гамма (IFN-gamma)
- угнетает. Растворимые низкоаффинные рецепторы FceRII
(CD23) в ассоциации с IL-4
способствуют дифференцировке В-лимфоцитов в IgE-синтезирующие
клетки, a
IFN-gamma
ингибирует этот процесс [7-9].
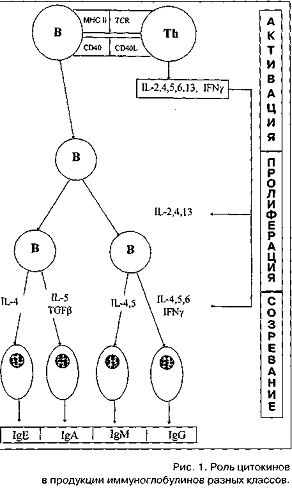
Рисунок 2. Роль цитокинов в
продукции иммуноглобулинов разных классов.
Дегрануляция тучных клеток и
базофилов наступает, когда две связанные с мембраной клеток молекулы IgE
соединяются с антигеном. После этого происходит выброс медиаторов воспаления
вследствие активации серинэстеразы, связанной с клеточной мембраной; притока
внутрь клетки ионов кальция; образования обладающих способностью к слиянию липидов
и последующего соединения периграну-лярной мембраны с плазматической мембраной;
снижения уровня внутриклеточного циклического 3',5'-аденозинмонофосфата;
сокращения микрофиламентов и агрегация микроканальцев. По мере развития этих
процессов гранула с медиатором сначала сообщается с внеклеточной средой при
помощи «цистерн», а затем, в конечном итоге, выталкивается из клетки [4,8,9].
Помимо участия в аллергических
реакциях I (немедленного)
типа, IgE также
принимает участие в защитном противогельминтном иммунитете (Рисунок 3), что
обусловлено существованием перекрестного связывания между IgE
и антигеном гельминтов. Последний, проникая через мембрану слизистой оболочки,
"садится" на тучные клетки и вызывает их дегрануляцию. Медиаторы
воспаления повышают проницаемость капилляров и слизистых оболочек, в результате
чего IgG и лейкоциты
выходят из кровотока. К гельминтам, покрытым IgG,
присоединяются эозинофилы, которые выбрасывают содержимое своих гранул и таким
образом убивают гельминтов [4, 9].
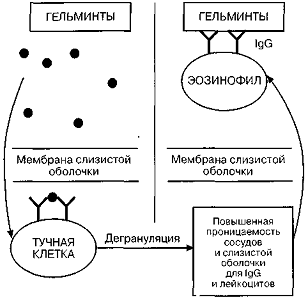
Рис. 3. Участие IgE
в защите против гельминтов
В норме IgE
составляет менее 0,001% от всего иммуноглобулина сыворотки крови. Его
концентрация в сыворотке крови взрослого человека составляет менее 0.05 г/л
(около 100 кЕ/л), что в 1000 раз и более ниже концентрации IgG
в сыворотке (около 10 г/л). Содержание IgE
в сыворотке крови здоровых людей находится в пределах указанных в Таблице 1.
Таблица 1.
Пределы содержания IgE
в сыворотке крови здоровых людей.
|
Возрастные
группы
|
Содержание
1дЕ(кЕ/л)
|
|
Новорожденные
|
0-2
|
|
Дети:
|
|
|
3
- 6 мес
|
|
1
год
|
8-20
|
|
5
лет
|
10-50
|
|
10
лет
|
15-60
|
|
Взрослые
|
20-100
|
IgE
можно обнаружить в организме человека уже на 11-й неделе внутриутробного
развития. Содержание IgE
в сыворотке крови возрастает постепенно с момента рождения человека до
подросткового возраста. В пожилом возрасте уровень IgE
может снижаться [4, 6. 9].
иммуноглобулин
сыворотка кровь гельминт эффекторный
Регуляция синтеза IgE
у человека
Таким образом, общая концепция
индукции и механизма регуляции продукции IgE,
разделяемая в настоящее время большинством исследователей, может быть
представлена в схематической форме в следующем виде (Рисунок 4).
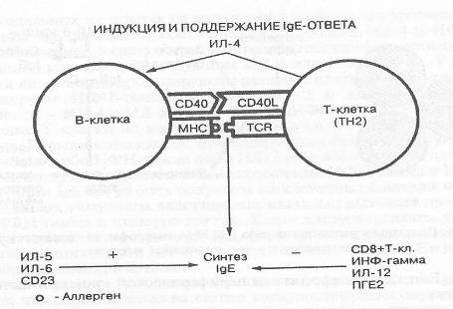
Рисунок 4. Регуляция синтеза IgE
у человека.
Антиген-представляющие клетки
после процессирования аллергена представляют его пептидные компоненты
(Т-клеточные эпитопы) на молекулах II
класса главного комплекса гистосовместимости Т-клеткам, которые распознают
соединение «пептид + главный комплекс гистосовместимости» (пептид+МНС) при
помощи Т-клеточного рецептора (TCR)
- «родственное» распознавание. При некоторых условиях в клеточный контакт
вовлекаются неспецифические мембранные компоненты («неродственное»
распознавание). Результатом этих взаимодействий является активация Т-клеток.
Избирательная стимуляция аллерген-специфических Т-клеток характеризуется
стимуляцией продукции ИЛ-4, а также других цитокинов. Сигнал, осуществляемый
ИЛ-4, ответственен за выбор изотипа (IgE)
иммуноглобулина, на который происходит переключение синтеза. Помимо MHC-TCR
взаимодействия и продукции цитокинов необходим другой сигнал, осуществляемый
взаимодействием молекулы CD40
на В-клетках и лиганда CD40L,
экспрессированного на Т-клетках. Этот второй сигнал, обязательный для перехода
синтеза иммуноглобулинов с одного изотопа на другой, и реализуется при действии
ИЛ-4 в переключение на синтез иммуноглобулина Е (Bonnefoy
et al.,
1996; Renz, 1995; Vercelli,
1995).
Разумеется, приведенная
концепция не является окончательной. Она постоянно дополняется, что со
временем, конечно, приведет к ее изменению.
Эффекторные
свойства IgE
Иммуноглобулин Е не обладает
комплементсвязывающей способностью по классическому пути. Лишь в очень больших
концентрациях IgE может
активировать комплемент (фиксация СЗ компонента комплемента) альтернативным
путем. Однако такая активность вряд ли может иметь какое-либо существенное
биологическое значение.
Наиболее характерным для IgE
биологическим свойством является его способность прочно фиксироваться на
определенных типах клеток - на тучных клетках и базофилах, что и определяет
сенсибилизацию организма, его органов и тканей. Это свойство закреплено за
определенными участками тяжелых цепей молекулы IgE.
Прогревание при 56°С, вызывающее потерю кожно-сенсибилизирующей активности
реагинной сыворотки, имеет точкой приложения своего действия как раз эти
участки тяжелых эпсилон-цепей. Восстановление дисульфидных связей
восстанавливающими агентами также нарушает цитотропное свойство IgE.
Таким образом, все четыре уникальные характеристики реагинов, упомянутые выше,
оказались свойственны иммуноглобулину Е и были объяснены после выяснения его
структуры и биологической активности отдельных фрагментов. Предполагали, что
прочное прикрепление молекулы IgE
к мембране клеток-мишеней обеспечивается фиксацией тяжелой цепи в области Сε3
и
Сε4
доменов.
В процессе фиксации молекулы иммуноглобулина его рецептор на мембране
клетки-мишени сначала распознается участком домена Сε4,
а
затем связывание осуществляется в области Сε3 или
Сε4.
На
заключительном этапе фиксации происходит закрепление молекулы иммуноглобулина Е
посредством вовлечения участка пептидной цепи в области между 330-м и 361-м
аминокислотными остатками в N-концевой
области Сε3 домена, благодаря
чему эпсилон-цепь прочно фиксируется в мембране тучной клетки или базофила.
Последующие исследования уточнили, что функция одного лишь С ε3
домена
является достаточной для фиксации IgE.
Так же как и у животных,
гомоцитотропная сенсибилизирующая активность сыворотки крови человека может
быть связана не только с IgE,
но и с антителами, принадлежащими к IgG.
Такая гетерогенность реагиновых антител человека, установленная в относительно
ранний период изучения их иммуноглобулиновой принадлежности (Адо и др., 1969),
в последующем была многократно подтверждена. Следует, однако, подчеркнуть, что
вклад этих антител в реагинную активность значительно меньше, чем IgE-антител.
Гомоцитотропные IgG-антитела
намного слабее фиксируются на клетках-мишенях, а потому легче и быстрее
удаляются с их поверхности отмывкой. Можно предположить, что аллергические
реакции, опосредуемые гомоцитотропными IgG-антителами,
явились в эволюции прототипом полноценной аллергической формы реактивности,
сформировавшейся после возникновения IgE
(Гущин, 1979).