Филогенез хрусталика и стекловидного тела
ФИЛОГЕНЕЗ ХРУСТАЛИКА И СТЕКЛОВИДНОГО
ТЕЛА
ВВЕДЕНИЕ
Зрение является важным физиологическим процессом, с помощью которого
человек и животные получают представление о величине, форме и цвете предметов,
их взаимном расположении и расстоянии и таким образом приобретают возможность
ориентироваться в окружающем пространстве.
Орган зрения, как и все другие органы чувств, в ходе филогенетического
развития претерпел сложную эволюцию, которая шла в направлении все большего и
лучшего приспособления глаза к восприятию окружающего мира.
Орган зрения в филогенезе проделал путь от отдельных эктодермального
происхождения светочувствительных клеток (у кишечнополостных) до сложно
устроенных парных глаз у млекопитающих. У позвоночных животных глаза
развиваются сложно: из боковых выростов мозга образуется светочувствительная
оболочка - сетчатка. Средняя и наружная оболочки глазного яблока, стекловидное
тело формируются из мезодермы (среднего зародышевого листка), хрусталик - из
эктодермы.
Хрусталик у рыб, амфибий округлый. Аккомодация достигается вследствие
перемещения хрусталика и сокращения особой передвигающей хрусталик мышцы. У
рептилий и птиц хрусталик способен не только перемещаться, но и изменять свою
кривизну. У млекопитающих хрусталик занимает постоянное место, аккомодация
осуществляется вследствие изменения кривизны хрусталика. Стекловидное тело,
имеющее вначале волокнистую структуру, постепенно становится прозрачным.
ФИЛОГЕНЕЗ ОРГАНА ЗРЕНИЯ
Простейшей формой зрения следует считать начало реакции на свет. Почти
все живущее чувствительно к свету. У растений световая реакция проявляется
гелиотропизмом (листья растений расположены перпендикулярно солнечным лучам,
головки цветущего подсолнечника в течение всего дня повернуты к солнцу).
У некоторых животных (минога) зрительные органы_не локализованы, покровы
их обладают общей раздражимостью по отношению к свету.
Большинство известных типов глаз возникли в Кембрийский период - около
530 млн лет назад, как датируются окаменелые останки трилобитов. Несомненно, что
возникновению глаза предшествовало эволюционное становление молекулярного
механизма фоторецепции и включение его в состав клеток, которые приобретали
сенсорную специализацию.
Собственно глазом следует называть орган, в котором связанные с нервной
системой фоторецепторные клетки соседствуют с клетками, содержащими
экранирующий пигмент. Кроме того, глаз должен содержать элементы, позволяющие
фокусировать лучи путем их преломления, дифракции или отражения.
Даже в пределах отдельных типов (например, стрекающие, кольчатые и
плоские черви, моллюски) можно наблюдать возникновение и исчезновение глаз,
усложнение или упрощение их структуры, очевидно, сообразно тем задачам, которые
ставит перед животным окружающая световая обстановка.
Следовательно, в филогенезе глаз как структура возникал неоднократно,
возможно более 40 раз. В пределах каждого структурного типа глаз может
встречаться большое число вариаций его конструкции и оптических приемов
формирования изображения, особенно у членистоногих.
Однако с меньшей вероятностью можно полагать независимое происхождение
генов, управляющих развитием основных молекулярных и клеточных элементов
органов зрения. Скорее всего, раз возникнув и претерпевая свою молекулярную
эволюцию, они многократно проявлялись в различных сочетаниях, формируя
многообразие клеточной и молекулярной основы зрительного процесса.
Поэтому понятен интерес исследователей клеточной и молекулярной биологии
зрения к стрекающим (тип Cnidaria), наиболее простым из современных многоклеточных животных, которые обладают
органами световой чувствительности. Среди них выделяются глаза-камеры медуз,
устройство которых подобно глазам брюхоногих и головоногих моллюсков, а также
хордовых животных.
ГЛАЗКИ И ГЛАЗА МЕДУЗ
Строение и клеточный состав. Многоклеточными светочувствительными
органами обладают свободноплавающие гидроидные (Hydrozoa), сцифоидные (Scyphozoa) и кубомедузы (Cubozoa), а также сидячие ставромедузы (Staurozoa). К этим органам относят глазные
(пигментные) пятна, пигментные ямки с линзами и без них, а также камерные глаза
с роговицей, хрусталиком и сетчаткой, состоящей из пигментных клеток и
цилиарных (ресничных) фоторецепторов. Глазные пятна и ямки еще собирательно
называют глазками, чтобы отличить от более сложно устроенных глаз, содержащих
специализированные оптические элементы. Фоторецепторные клетки медуз биполярные
по форме.
Их апикальный конец несет реснички, с характерным фибриллярным аппаратом
9+2. В свою очередь реснички, как и сама апикальная мембрана, могут
образовывать микровиллярные отростки. Аксон, берущий начало от базальной части
фоторецептора, образует контакты на вставочных нейронах. Вероятно,
взаимодействие осуществляется также и между рецепторными клетками сетчатки
гидро- и кубомедуз через синаптические контакты различных типов. Клетки,
содержащие гранулы экранирующего пигмента меланина, образуют микровиллярные
выросты, которые у некоторых видов достаточно велики и переплетаются с таковыми
фоторецепторов.
Сцифомедузы
одного и того же рода могут отличаться по признаку наличия глазков. Глазки
некоторых медуз отрядов Semaeostomeae и Rhizostomeae
<#"538906.files/image001.gif">
Рис.
1. Филогенез глаза (зрительные клетки дождевого червя)
Большой
интерес представляют активно плавающие многощетинковые кольчатые черви из
семейства Alciopidae, ведущие хищный образ жизни. У этих полихет глаз не только
отличается исключительно сложным строением и величиной, но и снабжен
аккомодационным устройством в виде специальных сократительных волокон,
способных передвигать хрусталик и тем самым менять фокусное расстояние. Это
единственный известный случай среди низших беспозвоночных: аккомодация глаза
встречается только у головоногих моллюсков и позвоночных. Возможно, у этих
червей в какой-то степени уже существует предметное зрение, что было бы
исключением, подтверждающим общее правило. Это относится и к свободно плавающим
хищным моллюскам Heteropoda, которые тоже обладают весьма сложно устроенными
глазами с приспособлением, заменяющим аккомодацию.
Что
же касается дождевого червя, то здесь обнаруживается чрезвычайно интересный
факт: у него нет не только сложно устроенных, но и вообще никаких специальных
органов светочувствительности. Вместе с тем ему свойствен четкий отрицательный
фототаксис. Функцию светоощущения выполняют рассеянные в коже
светочувствительные клетки. Это пример кожной светочувствительности низших
многоклеточных беспозвоночных. Кожная светочувствительность наблюдается и у
многих моллюсков, причем у двустворчатых это нередко единственная форма
фоторецепции. Эти моллюски реагируют как на освещение, так и на затемнение чаще
всего одинаковым образом - втягиванием выступающих из раковин частей тела или
запиранием раковины. Многие улитки реагируют на внезапное затемнение
сокращением «ноги», причем эта реакция сохраняется и после экстирпации глаз,
что опять-таки указывает на наличие кожной светочувствительности.
Реакция
дождевого червя на условия освещения состоит в том, что он уползает в зону
большего затемнения. Если же внезапно уменьшить интенсивность освещения, то
червь реагирует на это движениями бегства; в естественных условиях он вползает
в почву. Такая же реакция следует за внезапным освещением. Если же осветить
лишь определенный участок тела дождевого червя, то время этой реакции
сокращается пропорционально размеру освещаемой площади поверхности тела - в
наибольшей мере червь реагирует при освещении всей его поверхности.
Следовательно, реакция определяется градиентом раздражения освещенных и
неосвещенных частей тела. Аналогично реагируют на свет и другие кольчатые
черви.
Кольчатые
черви реагируют также на прикосновения, химические и термические раздражения,
силу тяжести, электрические раздражения, течение воды, а наземные формы (дождевые
черви) - на влажность среды. Однако принципиальных отличий от реакции на свет
нет: все эти реакции находятся на одном уровне и характеризуются тем, что
являются ответами на отдельные раздражители, на отдельные признаки, качества
предметов, но не на сами предметы как таковые. Так, например, много- и
малощетинковые черви проявляют четко выраженные таксисные реакции на тактильные
раздражения. Преобладают отрицательные реакции, но в ряде случаев наблюдаются и
положительные тигмотаксисы: прикосновения нервных сегментов животного к
субстрату влекут за собой прижимание к нему всем телом, что имеет, конечно,
большое значение при роющем образе жизни или жизни в трубках. Интересно, что у
дождевого червя соответствующие рецепторные образования представлены лишь отдельными
чувствительными клетками, разбросанными по всему телу, но более густо
располагающимися на его переднем конце. У многощетинковых же червей органами
осязания часто являются щупальца или щетинки.
Хорошо
развита и химическая чувствительность, причем в большинстве случаев наблюдается
отрицательный хемотаксис. При большой интенсивности химического воздействия
черви всегда реагируют отрицательно. С другой стороны, дождевой червь,
например, способен по химическим признакам выбирать разные виды листьев, что свидетельствует
о специализации в сенсорной сфере. У ряда кольчецов обнаружены расположенные
около ротового отверстия органы химической чувствительности в виде ямок.
Особенно это относится к пларающим видам, у которых имеется пара таких ямок,
выстланных мерцательным эпителием.
У
плоского червя орган зрения представляет собой чувствующую клетку,
продолжающуюся в нервный отросток и имеющую каемку из палочковидных окончаний.
Каемка покрыта особой пигментной клеткой. «Зрительные клетки» в глазу пиявки
объединены в группы по 5- 6 клеток (рис. 2).
В
глазах морской звезды обнаруживается начальная структура нейро-эпителия (рис.
3).
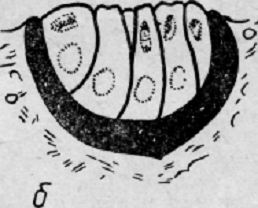
Рис.
2. Филогенез глаза (глаз пиявки)
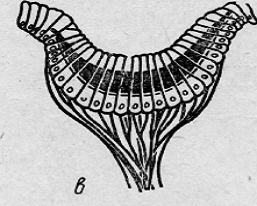
Рис.
3. Филогенез глаза (глаз морской звезды)
Его
световоспринимающие концы обращены к свету, нервные волокна собраны в широкий
рыхлый тяж, который можно считать примитивным нервом. Наружная часть глаза
имеет форму ямки, выстланной сверху покровным эпителием. Глаз кольчатых червей
еще более сложен (рис. 4).
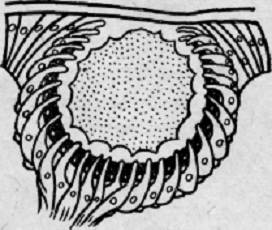
Рис.
4. Филогенез глаза (глаз кольчатого червя)
Он
имеет вид круглой полости, наполненной прозрачной массой - своеобразным стекловидным
телом. Между чувствительными клетками находятся клетки пигментного эпителия,
появляются вставочные клетки, что соответствует опорной, глиозной ткани
сетчатки высших животных. Если простейшие глаза реагируют только на свет и
изменение интенсивности света, то более развитые глаза способны формировать
образ.
Моллюск,
стоящий еще на достаточно низкой ступени развития, имеет глаз, который
напоминает глаз высших животных (рис. 5).
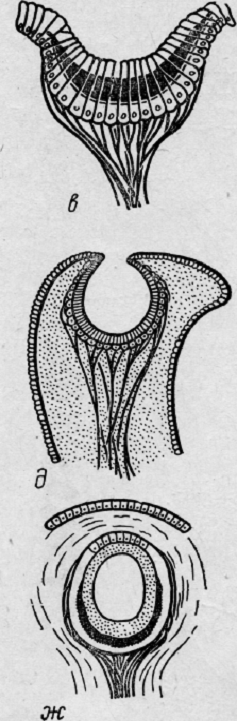
Рис.
5. Филогенез глаза (глаз моллюска)
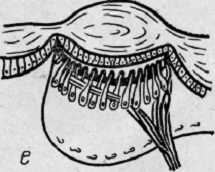
Рис.
6. Филогенез глаза (глаз скорпиона)
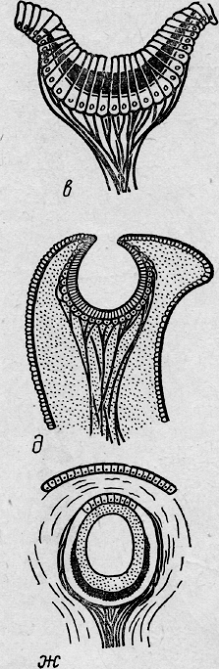
Рис.
7. Филогенез глаза (глаз улитки)
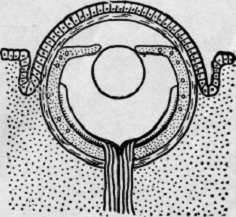
Рис.
8. Филогенез глаза ( глаз позвоночного)
Клетки
нейроэпителия обращены не к свету, не к центру глаза, а от света. Возникает тип
перевернутой сетчатки, что характеризует глаз высших животных. В глазу моллюска
уже есть подобие линзы. Фоторецепторы скрываются в углублениях, где они
защищены от яркого света, уменьшающего способность улавливать движущуюся тень.
Линза выполняет функцию прозрачной защитной мембраны. Постепенно начинает
совершенствоваться защитный аппарат глаза.
Сложно
организованные глаза часто бывают при простом мозге. У некоторых членистоногих
(включая насекомых) имеются сложные фасеточные глаза, содержащие свыше тысячи
фасеток. Такие глаза являются специальными детекторами движения и могут быть
чрезвычайно эффективными.
Глаз
человека по структуре представляет собой типичный глаз позвоночных, однако функциональные
различия его существенны.
РАЗВИТИЕ ГЛАЗА ЧЕЛОВЕКА
Глаз высших животных развивается из разных тканевых источников. Сетчатка
и зрительный нерв формируются из эктоневральной закладки центральной нервной
системы. На 2-й неделе эмбриональной жизни, когда мозговая трубка еще не
замкнута, на дорсальной поверхности медуллярной пластинки появляются два
углубления - глазные ямки. На вентральной поверхности им соответствует
выпячивание. При замыкании мозговой трубки ямки перемещаются, принимают боковое
направление. Эта стадия носит название первичного глазного пузыря (рис. 9).
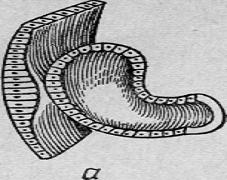
Рис. 9. Онтогенез глаза человека. ( первичный глазной пузырь)
В этой ранней стадии развития глаза полость мозга свободно переходит в
полость первичного глазного пузыря. Вершины глазных пузырей почти вплотную
подходят к эктодерме; их разделяет лишь узкий слой мезодермы. Такие соотношения
выявляются на 3-й неделе, когда длина всего зародыша 3 мм. С конца 4-й недели
развития возникает хрусталик (рис. 10).
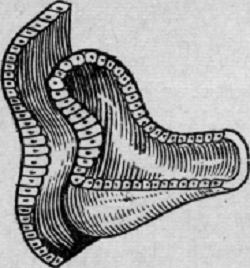
Рис. 10. Онтогенез глаза человека (закладка хрусталика в виде утолщения
эктодермы над первичным глазным пузырем)
Вначале он имеет вид утолщения покровной эктодермы в том месте, где
первичный глазной пузырь близко подходит к ней. В это же время первичный
глазной пузырь начинает превращаться во вторичный. Первичный пузырь растет
неравномерно, отмечается быстрый рост задних и боковых стенок первичного
глазного пузыря, в то время как рост передних и нижних стенок задерживается.
Быстрорастущие задние и боковые области обрастают передние и нижние части.
Однослойный первичный глазной пузырь на полой ножке превращается во вторичный
пузырь, состоящий из двух слоев - глазной бокал (рис. 11).
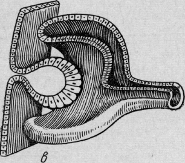
Рис. 11. Онтогенез глаза человека (вторичный глазной пузырь).
При образовании глазного бокала возникает зародышевая щель, которая
заполняется прилежащей мезодермой. Между зачатком хрусталика и внутренней
стенкой бокала остается небольшое количество мезенхимальных клеток, из которых
формируется первичное стекловидное тело (рис. 1).
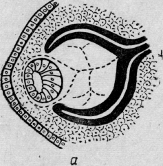
Рис. 12. Развитие глаза человека. Закладка первичного стекловидного тела
В этот период развития хрусталик занимает почти всю полость глазного
яблока. Очень интенсивно происходит размножение клеток, выстилающих внутреннюю
стенку хрусталикового пузырька. Постепенно вытягиваясь, они заполняют всю
полость пузырька. Край глазного бокала снизу начинает все больше ввертываться,
формируя вторичную зародышевую щель. Через эту щель проникает большое
количество мезенхимы, которая образует богатую сосудистую сеть стекловидного
тела. Вокруг хрусталика формируется сосудистая капсула. В возрасте 6 нед
зародышевая щель глаза и зрительного нерва закрывается, начинает
дифференцироваться ножка глазного бокала, образуется a. hyaloidea,
питающая стекловидное тело и хрусталик (рис. 13).
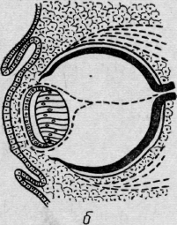
Рис. 13. Развитие глаза человека. Дифференцировка зрительного нерва.
Образование п. hyaloidea.
Наружный листок бокала в дальнейшем превращается в пигментный слой
сетчатки, из внутреннего же развивается собственно сетчатка. Края глазного
бокала, прорастая впереди хрусталика, образуют радужную и ресничную часть
сетчатки. Ножка, или стебелек, глазного бокала удлиняется, пронизывается
нервными волокнами, теряет просвет и превращается в зрительный нерв. Из
мезодермы, окружающей глазной бокал, очень рано начинают дифференцироваться
сосудистая оболочка и склера. В мезенхиме, которая прорастает между эктодермой
и хрусталиком, появляется щель - передняя камера. Мезенхима, лежащая перед
щелью, вместе с эпителием кожи превращается в роговицу, лежащая сзади -в
радужку. К этому времени начинается постепенное запустение сосудов
стекловидного тела. Сосудистая капсула хрусталика атрофируется. Внутри
хрусталика образуется плотное ядро (зародышевое ядро хрусталика), объем
хрусталика уменьшается. Стекловидное тело приобретает прозрачность (рис. 14).
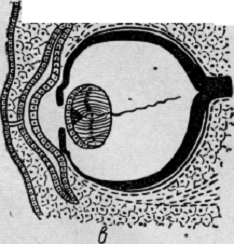
Рис. 14. Развитие глаза человека. Дифференцировка оболочек глаза.
Веки развиваются из кожных складок. Они закладываются кверху и книзу от
глазного бокала, растут по направлению друг к другу и спаиваются вместе своим
эпителиальным покровом. Спайка эта исчезает к 7 мес развития.
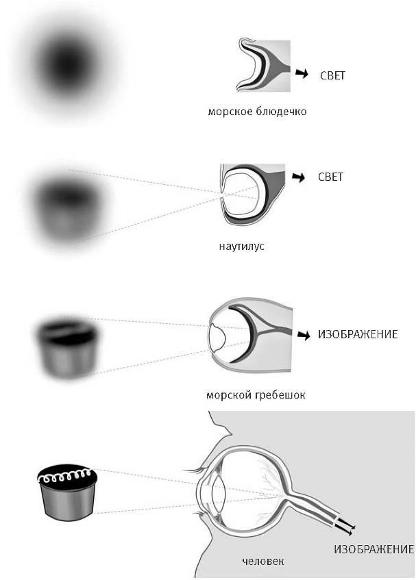
Рисунок 15. Сравнительная характеристика
Слезная железа возникает на 3-м месяце, слезный канал открывается в
носовую полость на 5-м месяце. К моменту рождения ребенка весь сложный цикл
развития глаза не всегда оказывается полностью завершенным. Обратное развитие
элементов зрачковой перепонки, сосудов стекловидного тела и хрусталика может
происходить и в первые недели после рождения.
Хрусталик (lens crystallina) развивается из эктодермы. Это
исключительно эпителиальное образование. Он изолирован от остальных оболочек
глаза капсулой, не содержит нервов, сосудов и других каких-либо мезодермальных
клеток. В связи с этим в хрусталике не могут возникать воспалительные процессы.
Хрусталиковый карман, возникший из инвагинированной поверхностной
эктодермы и вместившийся в вогнутость глазного бокала, полностью
отшнуровывается от поверхностной эктодермы. В этой стадии развития (на
четвертой неделе) хрусталиковый карман представляет собой яйцевидный пузырь, в
котором распознается передняя стенка, образованная низкими кубическими
клетками, и задняя стенка, клеточный слой которой состоит из более высоких
цилиндрических клеток. По краям (по будущему экватору хрусталика) оба слоя
сливаются между собой, замыкая тем самым полость.
При дальнейшем развитии из кубических клеток передней хрусталиковой
стенки образуется передний эпителий хрусталика, причем клетки здесь остаются
сравнительно невысокими. Наоборот, клетки задней хрусталиковой стенки
преобразуются в хрусталиковые волокна.
Исходные клетки начинают удлиняться и расти по направлению к передней
стенке, благодаря чему вся полость хрусталикового пузыря постепенно исчезает.
Хрусталиковые волокна при этом теряют способность делиться и размножаться, их
ядра дегенерируют и исчезают.
Новые хрусталиковые волокна вырастают лишь из экваториальной приростковой
зоны, прилегая затем к хрусталиковому ядру, которое возникает из первоначальных
клеток заднего слоя, благодаря чему происходит увеличение объема хрусталика.
Вновь возникшие поверхностные волокна не только врастают под передний
хрусталиковый эпителий, но и обрастают заднюю поверхность хрусталика, огибая
его при этом по экватору. При этом всегда сохраняется определенное отношение
между длиной передней и задней частей новых хрусталиковых волокон.
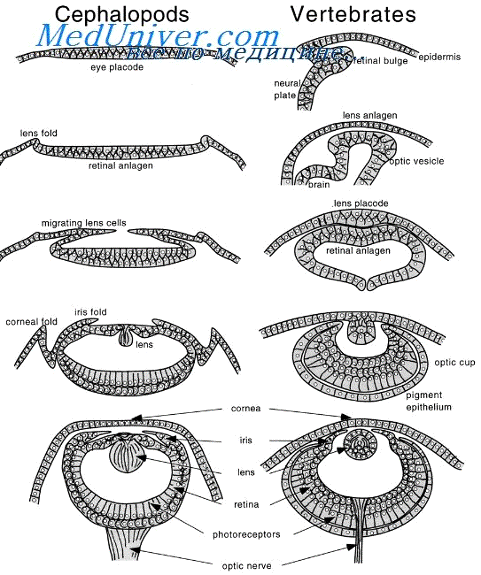
Рис. 16 Этапы формирования хрусталика
Чем длиннее передняя часть волокна, тем короче ее задняя часть и
наоборот. Волокна, которые таким образом возникают по всей окружности экватора,
послойно налегают друг на друга, и, переплетаясь на передней и задней
поверхности хрусталика, создают характерные звездчатые хрусталиковые швы,
имеющие сначала только три или четыре плеча, впоследствии и большее количество
плеч. Вокруг всего хрусталика образуется бесструктурная капсула, являющаяся
продуктом жизнедеятельности клеток стенок хрусталикового пузыря (сумка
хрусталика - capsula lentis).
У взрослого человека хрусталик представляет собой прозрачное, слегка
желтоватое, сильно преломляющее свет тело, имеющее форму двояковыпуклой линзы.
По силе преломления хрусталик является второй средой (после роговицы)
оптической системы глаза. Его преломляющая сила в среднем 18,0 D. Расположен хрусталик между радужкой
и стекловидным телом, в углублении передней поверхности последнего. Удерживают
его в этом положении волокна поддерживющей связки (zonula ciliaris), которые другим своим концом
прикрепляются по внутренней поверхности цилиарного тела.
Хрусталик состоит из хрусталиковых волокон, составляющих вещество
хрусталика, и сумки-капсулы. Консистенция хрусталика в молодые годы мягкая. С
возрастом увеличивается плотность центральной его части, поэтому принято
выделять кopу хрусталика и ядро хрусталика. В
хрусталике различают экватор и два полюса - передний и задний (рис. 17).
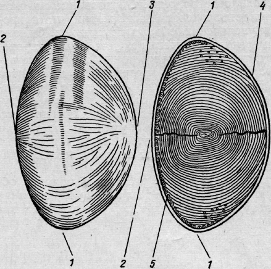
Рис. 17. Хрусталик.
экватор; 2 - передний полюс; 3 - задний полюс; 4 - капсула; 5 - эпителий.
Условно по экватору хрусталик делят на переднюю и заднюю поверхности.
Линия, соединяющая передний и задний полюса, называется осью хрусталика.
Диаметр хрусталика 9- 10 мм. Передне-задний его размер 3,5 мм. Передняя
поверхность хрусталика менее выпуклая, чем задняя.
Гистологически хрусталик состоит из капсулы, эпителия капсулы и волокон. Эпителий
покрывает лишь внутреннюю поверхность передней капсулы, поэтому носит название
эпителия передней сумки. Клетки его имеют шестиугольную форму. У экватора
клетки приобретают вытянутую форму и превращаются в хрусталиковое волокно.
Образование волокон совершается в течение всей жизни, что приводит к увеличению
объема хрусталика. Однако чрезмерного увеличения хрусталика не происходит, так
как центральные, более старые волокна теряют воду, оплотневают, становятся уже
и постепенно в центре образуют компактное ядро. Это явление склерозирования
следует расценивать как физиологический процесс, который приводит лишь к
уменьшению объема аккомодации (см. раздел «Аккомодация»), но практически не
снижает прозрачности хрусталика.
Хрусталик вместе с ресничным пояском образует цилиохрусталиковую
диафрагму, которая делит полость глаза на две неравные части: меньшую -
переднюю и большую- заднюю.
Стекловидное тело
Стекловидное тело (corpus vitreum) является
частью оптической системы глаза. Оно выполняет полость глазного яблока, за
исключением передней и задней камер глаза, и таким образом способствует
сохранению его тургора и формы. По мнению ряда исследователей, стекловидное
тело в известной степени обладает амортизирующими свойствами, поскольку его
движения сначала являются равномерно-ускоренными, а затем
равномерно-замедленными. Объем стекловидного тела взрослого человека 4 мл. Оно
состоит из плотного остова и жидкости, причем на долю воды приходится около 99%
всего состава стекловидного тела. Тем не менее вязкость стекловидного тела в
несколько десятков раз выше вязкости воды. Вязкость стекловидного тела,
являющегося гелеобразной средой, зависит от содержания в его остове особых
белков - витрозина и муцина. С мукопротеидами связана гиалуроновая кислота,
играющая важную роль в поддержании тургора глаза.
Стекловидное тело, или corpus vitreum, заполняет полость глазного яблока
между задней поверхностью хрусталика и сетчатой оболочкой. Оно в течение
второго месяца закладывается сначала в виде тонких плазматических мостиков между
внутренним листком глазного бокала и задней хрусталиковой поверхностью.
Стекловидное тело берет, по-видимому, начало из обоих этих эпителиальных
слоев (из сосудистой оболочки и из хрусталика). Между упомянутыми волокнистыми
мостиками откладывается студенистая, стекловидно прозрачная масса. В
формировании окончательного состава стекловидного тела, по всей вероятности,
принимает участие и мезенхимная ткань, которая попадает сюда еще до полного
замыкания эмбриональной глазной щели, (сосудистой щели - fissura chorioidea), в
глазном бокале.
Рис. 18 Стекловидное тело
Посередине стекловидной ткани проходит стекловидная артерия (arteria
hyaloidea), после обратного развития которой в глазу взрослого человека
сохраняется лишь едва заметный каналец - канал стекловидного тела - canalis
hyaloideus (Cloqueti).
Средний васкуляризированный слой (tunica vasculosa) и наружный фиброзный
слой (tunica fibrosa) глазного яблока развиваются из сгущенной мезенхимы,
находящейся вокруг закладки глазного бокала, который приблизительно на седьмой
неделе делится на два слоя.
Наружный слой состоит из сгущенной, уплотнившейся мезенхимной ткани, он
перестраивается в плотную фиброзную ткань склеры белочной оболочки глазного
яблока и роговой оболочки. Слой склеры в области зрительного нерва связан
непосредственно с его соединительнотканной оболочкой; по ходу нерва склера
незаметно переходит в твердую оболочку мозга.
В передней части глаза эта ткань дифференцируется на прозрачную роговую
оболочку. Часть эдидермиса (наружной эктодермы), из которого уже до этого в
данном месте отделился хрусталик, вновь восстанавливается и несколько позже
дифференцируется на равномерно наслоенный (пятислойный) плоский эпителий
(передний роговой эпителий). Кнаружи от рогового слоя этот эпителий
продолжается в конъюнктивальную выстилку.
Между передним роговым эпителием и пузырем хрусталика из окружающей среды
врастает мезенхима, которая затем дает начало возникновению как узкого
внутреннего слоя, так и более широкого наружного слоя. Из внутреннего слоя,
прилежащего спереди к сумке хрусталика, образуется уже упомянутая зрачковая
мембрана (сосудистая оболочка хрусталика) - membrana pupillaris (tunica
vasculosa Ientis). Наружный, более широкий слой мезенхимы прикладывается к переднему
эпителию роговой оболочки, и из нее развивается собственное вещество, или
строма роговины (substantia propria corneae), задняя пограничная, или
десцеметова, мембрана (membrana Descemeti) и роговой, корнеальный, эндотелий
(задний эпителий роговой оболочки).
Из этой мезенхимы возникает также и соединительнотканный компонент
конъюнктивы глазного яблока и передней части склеры.
По химическому составу стекловидное тело очень сходно с камерной влагой,
а также со спинномозговой жидкостью.
Для понимания особенностей строения стекловидного тела и патологических
изменений в нем необходимо иметь представления об этапах его развития.
Первичное стекловидное тело является мезодермальным образованием и весьма
далеко от окончательного своего вида - прозрачного геля. Вторичное стекловидное
тело состоит из мезодермы и эктодермы. В этот период начинает формироваться
волокнистый остов стекловидного тела (из сетчатки и цилиарного тела).
Сформированное стекловидное тело (третий период) остается постоянной
средой глаза. При потере оно не регенерирует и замещается внутриглазной
жидкостью.
Стекловидное тело прикрепляется к окружающим его отделам глаза в
нескольких местах.
Главное место прикрепления называют о с новой, или базисом, стекловидного
тела. Основа представляет собой кольцо, выступающее несколько кпереди от
зубчатой линии. В области базиса стекловидное тело прочно связано с цилиарным
эпителием. Эта связь настолько прочна, что при отделении стекловидного тела от
основы в изолированном глазу вместе с ним отрываются эпителиальные части
цилиарных отростков, оставаясь прикрепленными к стекловидному телу. Второе по
прочности место прикрепления стекловидного тела - к задней капсуле хрусталика -
называется гиалоидохрусталиковой связкой, имеющей важное клиническое значение.
При интракапсулярной экстракции катаракты целость ее может нарушиться, что
ведет к выпадению стекловидного тела.
Третье заметное место прикрепления стекловидного тела приходится на
область диска зрительного нерва и по размерам соответствует примерно площади
диска зрительного нерва. Это место прикрепления- наименее прочное из
перечисленных трех.
Существуют и более слабые прикрепления стекловидного тела в области
экватора глазного яблока.
Большинство исследователей считают, что стекловидное тело особой
пограничной оболочкой не обладает. Большая плотность переднего и заднего
пограничных слоев зависит от имеющихся здесь несколько более густо
расположенных нитей остова стекловидного тела. При электронной микроскопии
установлено, что стекловидное тело имеет фибриллярную структуру. Фибриллы имеют
величину около 25 нм. Достаточно изучена топография гиалоидного, или клокетова,
канала, через который в эмбриональном периоде от диска зрительного нерва к
задней поверхности хрусталика проходит артерия стекловидного тела (a. hyaloidea). Ко времени рождения a. hyaloidea исчезает, а гиалоидный канал
сохраняется в виде узкой трубочки. Канал имеет извилистый S-образный ход. В середине
стекловидного тела гиалоидный канал поднимается кверху, а в заднем отделе имеет
тенденцию располагаться горизонтально.
Водянистая влага, хрусталик, стекловидное тело вместе с роговицей
образуют преломляющие среды глаза, обеспечивающие отчетливое изображение на
сетчатке. Заключенные в замкнутую со всех сторон капсулу глаза водянистая влага
и стекловидное тело оказывают ют известную степень напряжения, обусловливают
тонус глаза, внутриглазное давление (tensio oculi).
ЛИТЕРАТУРА
филогенез хрусталик
стекловидное тело
1. А.Б. Ходжаян, Н.Н. Федоренко, Л.А. Краснова О некоторых
морфо-функциональных преобразованиях нервной, эндокринной систем и органов
чувств в филогенезе беспозвоночных и позвоночных. // Учебно-методическое
пособие для студентов 1-го курса СтГМА. Ставрополь 2007. - 88 с.
2. Алексеева Т. И. Адаптивные процессы в популяциях
человека. М., 1986.
. Алмазов И.В., Сутулов Л.С. - Атлас по гистологии и
эмбриологии
. Афанасьев Ю.И., Юрина Н.А. - Гистология, цитология и
эмбриология. М. 2002
. Афанасьев Ю.И., Юрина Н.А., ред. Гистология,
цитология и эмбриология. 5-е изд. М.: «Медицина», 2002, 737 с.
. Бигон М., Харпер Дж., Таунсенд К. Экология. Особи,
популяции, сообщества. М., 1989. Т. 1, 2.
. Быков Б.Л. Частная гистология человека. 2-е изд.
СПб.: «Сотис», 2002, 298 с.
. Валькович Э.И., Батюто Т.Д., Кожухарь В.Г.,
Столярова М.В., Чурилова Н.И., Раковщик А.Л., Игнатенко Ю.В., Зажицкая Е.
Материалы к лекциям по гистологии и эмбриологии. Кафедра гистологии и
эмбриологии СПбГПМА, 2006.
. Заварзин А.А. Основы сравнительной гистологии. Л.,
1985.
. Заварзин А.А. Сравнительная гистология. Изд.
Санкт-Петербургского университета, 2000, 520 с.
. Казначеев В. П. Учение В. И. Вернадского о биосфере
и ноосфере. Новосибирск,1989.
. Костюк П.Г. Физиология центральной нервной системы.
М.:Изд-во МГУ, 1997.
. Крумбигель Г., Вольтер X. Ископаемые. М., 1988.
. Курс зоологии / Под ред. Б. С. Матвеева. М., 1961.
Т. 1, 2.
. Левонтин Р. Генетические основы эволюции. М., 1978.
. Левушкин С.И., Шилов И.А. Общая зоология. М., 1994.
. Майр Э. Популяции, виды и эволюция. М., 1974.
. Н.В. Воронова, Н.М. Климова, А.М. Менджерицкий.
Анатомия Центральной Нервной Системы. Москва 2006
. Рева Г.В., Рева И.В., Мельникова Н.В., Кислякова
И.С. КЛЕТКИ СТЕКЛОВИДНОГО ТЕЛА ГЛАЗА ЧЕЛОВЕКА // Успехи современного
естествознания. - 2009. - № 1 - С. 51-53
. Рогинский Я.Я., Левин М.Г. Антропология. М., 1978.
. Ромер А., Пирсоне Т. Анатомия позвоночных. М., 1992.
Т. 1, 2.
. Рэфф Р., Кофмен Т. Эмбрионы, гены и эволюция. М.,
1986.
. Тимофеев-Ресовский Н.В., Воронцов Н.Н., Яблоков А.В.
Краткий очерк теории эволюции. М., 1987.
. Улумбеков Э.Г., Челышев Ю.А. (ред.), Бойчук Н.В. и
др. Гистология. Учебник для вузов. ГЭОТАР - МЕД-2001
. Фоули Р. Еще один неповторимый вид. Экологические
аспекты эволюции человека. М., 1990.
. Шульговский В.В. Основы нейрофизиологии. М.: Аспект
Пресс, 2000.
. Эволюционное учение. Яблоков А.В., Юсуфов А.Г. М.:
Высшая школа, 2006. - 310 с