Типы деления клетки. Мегаспорогенез
Типы деления клетки.
Биологическое значение редукционного деления
деление клетка плод
мегаспоры капустные
Способность к делению - важнейшее свойство клеток. Без деления невозможно представить
себе увеличение числа одно клеточных существ, развитие сложного многоклеточного
организма из одной оплодотворенной яйцеклетки, возобновление клеток, тканей и
даже органов, утраченных в процессе жизнедеятельности организма. Деление клеток
осуществляется поэтапно. На каждом этапе деления происходят определенные
процессы. Они приводят к удвоению генетического материала (синтезу ДНК) и его
распределению между дочерними клетками. Период жизни клетки от одного деления
до следующего называется клеточным циклом.
Амитоз
Амитоз, или прямое деление, - это
деление интерфазного ядра путем перетяжки без образования веретена деления
(хромосомы в световом микроскопе вообще неразличимы). Такое деление встречается
у одноклеточных организмов (например, амитозом делятся полиплоидные большие
ядра инфузорий), а также в некоторых высокоспециализированных клетках растений
и животных с ослабленной физиологической активностью, дегенерирующих,
обреченных на гибель, либо при различных патологических процессах, таких как
злокачественный рост, воспаление и т. п. Амитоз можно наблюдать в тканях растущего
клубня картофеля, эндосперме, стенках завязи пестика и паренхиме черешков
листьев. Такой тип деления характерен для клеток печени, хрящевых клеток,
роговицы глаза. Очень часто при амитозе наблюдается только деление ядра, в этом
случае могут возникнуть двух- и многоядерные клетки. Если же за делением ядра
следует деление цитоплазмы, то распределение клеточных компонентов, как и ДНК,
осуществляется произвольно. Амитоз в отличие от митоза является самым
экономичным способом деления, так как энергетические затраты при этом весьма
незначительны. К амитозу близко клеточное деление у прокариот. Бактериальная
клетка содержит только одну, чаще всего кольцевую молекулу ДНК, прикрепленную к
клеточной мембране. Перед делением клетки ДНК реплицируется, и образуются две
идентичные молекулы ДНК, каждая из которых также прикреплена к клеточной
мембране. При делении клетки клеточная мембрана врастает между этими двумя
молекулами ДНК, так что в конечном итоге в каждой дочерней клетке, оказывается,
по одной идентичной молекуле ДНК. Такой процесс лучил название прямого
бинарного деления.
Подготовка к делению. Эукариотические организмы, состоящие из клеток, имеющих ядра,
начинают подготовку к делению на определенном этапе клеточного цикла, в
интерфазе. Именно в период интерфазы в клетке происходит процесс биосинтеза
белка, удваиваются все важнейшие структуры клетки. Вдоль исходной хромосомы из
имеющихся в клетке химических соединений синтезируется ее точная копия,
удваивается молекула ДНК. Удвоенная хромосома состоит из двух полови нок-
хроматид. Каждая из хроматид содержит одну молекулу ДНК. Интерфаза в клетках
растений и животных в среднем продолжается 10-20 ч. Затем наступает процесс
деления клетки - митоз.
Митоз
Митоз (от греч. Mitos- нить)
непрямое деление, - основной способ деления эукариотических клеток. Митоз - это
деление ядра, которое приводит к образованию двух дочерних ядер, в каждом из
которых имеется точно такой же набор хромосом, что и в родительском ядре. Вслед
за делением ядра обычно следует деление самой клетки, поэтому часто термином -
«митоз» обозначают деление клетки целиком. Митоз впервые наблюдали в спорах
папоротников, хвощей плаунов Г. Э. Руссов, преподаватель Дерптского универси
тета в 1872 г. и русский ученый И. Д. Чистяков в 1874 г. Де тальные исследования
поведения хромосом в митозе были выполнены немецким ботаником Э. Страсбургером
в 1876- 1879 гг. на растениях и немецким гистологом В. Флеммингом в 1882 г. на
животных. Митоз представляет собой непрерывный процесс, но для удобства
изучения биологи делят его на четыре стадии в зависимости оттого, как выглядят
в это время хромосомы в световом микроскопе. В митозе выделяют профазу,
метафазу, анафазу и телофазу. В профазе происходит укорочение и утолщение
хромосом вследствие их спирализации. В это время хромосомы двойные состоят из
двух сестринских хроматид, связанных между собой. Одновременно со спирализацией
хромосом исчезает ядрышко и фрагментируется (распадается на отдельные цистерны)
ядерная оболочка. После распада ядерной оболочки хромосомы свободно и беспорядочно
лежат в цитоплазме. В профазе центриоли (в тех клетках, где они есть)
расходятся к полюсам клетки. В конце профазы начинает образовываться веретено
деления, которое формируется из микротрубочек путем полимеризации белковых
субъединиц.
В метафазе завершается образование
веретена деления, которое состоит из микротрубочек двух типов: хромосомных,
которые связываются с центромерами хромосом, и центросомных (полюсных), которые
тянутся от полюса к полюсу клетки. Каждая двойная хромосома прикрепляется к
микротрубочкам веретена деления. Хромосомы как бы выталкиваются микротрубочками
в область экватора клетки, т. е. располагаются равном расстоянии от полюсов.
Они лежат в одной плоскости и образуют так называемую экваториальную,
или метафазную пластинку. В метафазе отчетливо видно двойное строение
хромосом, соединенных только в области центромеры. В этот период легко
подсчитывать число хромосом, изучать их морфологические особенности. В анафазе
дочерние хромосомы с помощью микротрубочек веретена деления растягиваются к
полюсам клетки. Во время движения дочерние хромосомы несколько изгибаются на
подобие шпильки, концы которой повернуты в сторону экватора клетки. Таким
образом, в анафазе хроматиды удвоенные в интерфазе хромосом расходятся к
полюсам клетки. В этот момент в клетке находятся два диплоидных набора
хромосом. В телофазе происходят процессы, обратные тем, которые наблюдаются в
профазе: начинается деспирализация (раскручивание) хромосом, они набухают и
становятся плохо видимыми под микроскопом. Вокруг хромосом у каждого полюса из
мембранных структур цитоплазмы формируется ядерная оболочка, в ядрах возникают
ядрышки.
Разрушается веретено деления. На
стадии телофазы происходит разделение цитоплазмы (цитотомия) с образованием
двух клеток. В клетках животных плазматическая мембрана начинает впячиваться
внутрь области, где располагался экватор веретена. В результате впячивания
образуется непрерывная борозда, опоясывающая клетку по экватору и постепенно
разделяющая одну клетку на две. В клетках растений в области экватора из
остатков нитей веретена деления возникает бочковидное образование - фрагмопласт.
В эту область со стороны полюсов клетки устремляются многочисленные пузырьки
комплекса Гольджи, которые сливаются друг с другом. Содержимое пузырьков
образует клеточную пластинку, которая делит клетку на две дочерние, а мембрана
пузырьков Гольджи образует недостающие цитоплазматические мембраны этих клеток.
Впоследствии на клеточную пластинку со стороны каждой из дочерних клеток
откладываются элементы клеточных оболочек. В результате митоза из одной клетки
возникают две дочерние с тем же набором хромосом, что и в материнской клетке.
Биологическое значение
митоза состоит, таким образом, в строго
одинаковом распределении между дочерними клетками материальных носителей
наследственности - молекул ДНК, входящих в состав хромосом. Благодаря
равномерному распределению реплицированных хромосом происходит восстановление
органов и тканей после повреждения. Митотическое деление клеток является также
цитологического размножения организмов.
Мейоз - это особый способ деления клеток, в результат которого
происходит редукция (уменьшение) числа хромосом вдвое. Впервые он был описан В.
Флеммингом в 1882 г. у животных и Э. Страсбургером в 1888 г. у растений. С
помощью мейоза образуются гаметы. В результате редукции споры и половые клетки
хромосомного набора в каждую гаплоидную спору и гамету по одной хромосоме из
каждой пары хромосом, имеющихся в данной диплоидной клетке. В ходе дальнейшего
процесса оплодотворения (слияния гамет) организм нового поколения получит опять
диплоидный набор хромосом, т. е. кариотип организмов данного вида в ряду
поколений остается постоянным.
Таким образом, важнейшее
значение мейоза заключается в обеспечении постоянства кариотипа в ряду
поколений организмов данного вида при половом размножении.
Редукционное деление является, по
сути, механизмом препятствующим непрерывному увеличению числа хромосом при
слиянии гамет, без него при половом размножении число хромосом удваивалось бы в
каждом новом поколении. Иными словами, б л а год ар я мейозу
поддерживает определенное и постоянное число хромосом во всех поколениях любого
вида растений, животных и грибов. Другое важное значение мейоза заключается
в обеспечении чрезвычайного разнообразия генетического состава гамет, как в
результате кроссинговера, так и в результате различного сочетания отцовских и
материнских хромосом при их независимом расхождении в анафазе I мейоза,
что обеспечивает появление разнообразного и разнокачественного потомства при
половом размножении организмов.
Ассимиляционная,
поглощающая паренхимы, их строение, функции, местоположение в органах растений.
Приведите примеры и рисунки
Паренхима - (от греч . parenchyma,
букв. - налитое рядом), 1) у растений - основная ткань из клеток более или
менее одинакового размера; осуществляет ассимиляцию, выделение и другие
функции. Разновидности паренхимы: поглощающая, ассимиляционная (хлоренхима),
запасающая, воздухоносная (аэренхима). 2) У животных паренхимой называют
главную функциональную ткань печени, селезенки, легкого и некоторых других
органов. ;
Ткани, основной функцией которых
является работа ассимиляции, то есть фотосинтез, объединяют в систему
ассимиляционных тканей.
У высших растений
они имеют обычно зеленую окраску. Поэтому могут быть названы зеленой паренхимой
или хлорофиллоносной паренхимой (короче ≈ хлоренхимой).
Ассимиляционная ткань устроена
достаточно просто и состоит из однородных тонкостенных клеток.
Хлоропласты в клетках хлоренхимы
обычно расположены в один ряд в постенном слое цитоплазмы. Центральная часть
полости клетки занята крупной вакуолью.
В соответствии с тем, что работа
ассимиляции происходит за счет солнечной энергии, хлоренхима располагается в
местах, наиболее доступных свету: она находится в надземной части растений непосредственно
под кожицей листьев и стеблей.
Нередко хлоренхима дифференцирована
на столбчатую (палисадную) и губчатую ткань. Палисадная
ткань обычно состоит из удлиненных клеток цилиндрической формы, расположенных
перпендикулярно к поверхности органа. Межклеточники в палисадной ткани развиты
слабо. Палисадная ткань содержит большое количество хлоропластов, здесь
происходят световые реакции фотосинтеза.
Губчатая ткань
построена из округлых или неопределенной формы клеток, образующих рыхлую
сложную сетчатую систему. Межклетники хорошо развиты. Здесь протекает газообмен
и темновая стадия фотосинтеза.
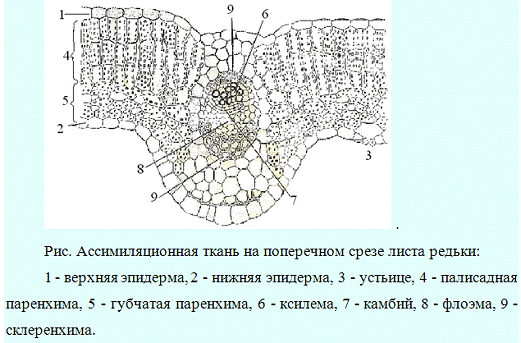
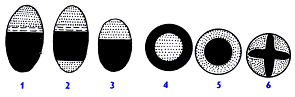
Рис. Типы
проводящих пучков
Схематичное
изображение строения различных типов проводящих пучков на поперечном их срезе
(ксилема зачернена, флоэма показана точками, а камбий - продольными штрихами).
1 -
открытый коллатеральный, 2 - открытый биколлатеральный, 3 - закрытый
коллатеральный, 4 - концентрический закрытый центрофлоэмный, 5 - концентрический
закрытый центроксилемный, 6 - закрытый радиальный проводящий пучок.
Обособленные тяжи
проводящей системы, состоящие чаще из ксилемы и флоэмы
<#"536938.files/image003.gif">
Рис.
Пшеница - род
травянистых, в основном однолетних, растений семейства Злаки, или Мятликовые
<#"536938.files/image004.gif">
Рис.
Итак, оба семени покрыты покровным
слоем, имеют эндосперм, зародыш, состоящий семядолей, стебелька, почечки,
корешка. Строение семени гороха и пшеницы отличается наличием количества
семядолей. У гороха -2, пшеницы-1.
Таблица. Химический состав семян
гороха и пшеницы
|
микроэлементы
|
горох
|
пшеница
|
|
Холин <#"536938.files/image005.gif">
Рис. Цикл развития сосны
обыкновенной: а - ветка с шишками; б- женская шишка в разрезе; в - семенная
чешуя с семязачатками; г - семязачаток в разрезе; д -мужская шишка в разрезе; е
- пыльца; ж - семенная чешуя с семенами; 1 - мужская шишка; 2 - молодая женская
шишка; 3 - шишка с семенами; 4 - шишка после высыпания семян; 5 - пыльцевход; 6
- покров; 7 - пыльцевая трубка со спермиями; 8 - архегоний с яйцеклеткой; 9 -
эндосперм.
Характеристика семейства
Капустные ( Крестоцветные). Укажите наиболее распространенные овощные, сорные и
дикорастущие растения этого семейства (не менее 15 видов)
Семейство крестоцветные
(Brassicaceae или Cruciferae)
Крестоцветные настолько близки к
каперсовым, что между ними не всегда легко провести границу. Некоторые роды,
например род диптеригиум (Dipterygium), одними ботаниками включается в
семейство каперсовые, а другими - в крестоцветные. В семействе насчитывается до
380 родов и. около 3200 видов. Расселены они по земному шару крайне
неравномерно. В основном сконцентрированы в умеренной зоне северного полушария,
главным образом в Старом Свете. В тропиках представлены единичными родами,
приуроченными к горным областям, встречаются там также в интродукции и как
сорняки. Небольшое число крестоцветных, произрастающих в южном полушарии, имеет
узколокальную приуроченность.
Крестоцветные успешно
приспосабливаются к самым разнообразным местообитаниям. Одни из них приурочены
к крайним условиям высокогорий, достигая рубежей растительности (4500-5700 м
над уровнем моря), где вместе с лишайниками являются пионерами растительного
покрова; другие произрастают по морским побережьям; одни в своем
распространении продвигаются далеко на север и характерны для арктических
областей; другие являются обитателями пустынь, полупустынь и степей. Широко
представлены крестоцветные также в лесах, среди степной растительности, на
увлажненных местах и даже в воде, но все же определенно преобладают среди них
растения засушливых и сухих местообитаний. Однако, несмотря на такую высокую
пластичность в приспособлении к условиям среды, наблюдается относительно
небольшое разнообразие жизненных форм. Большинство крестоцветных - однолетние
или многолетние травы, есть и полукустарнички, у которых одревесневает нижняя
часть стебля. Кустарники представлены единичными, преимущественно африканскими
и макаронезийскими видами, такими, как, например, катран кустарниковый (Crambe
fruticosa) на острове Мадейра, достигающий в высоту 2 м, виды рода
синапидендрон (Sinapidendron, Макаронезия), гелиофила сизая (Heliophila glauca
- Капская область) или фолейола Биллота (Foleyola billotii - Сахара),
достигающими в высоту до 1,5-2 м. Такие виды, как гелиофила лазящая (H.
scandens), и виды южноамериканского рода кремолобус (Cremolobus) габитуально
сближаются с лианами. Многие из высокогорных видов имеют подушкообразную форму,
способствующую задержанию тепла. Листья крестоцветных очередные, причем нижние
часто образуют прикорневую розетку. У некоторых видов наблюдается гетерофиллия.
Например, у клоповника пронзеннолистного (Lepidium perfoliatum) розеточные
листья рассечены на узкие линейные доли, тогда как стеблевые цельные, округлые,
стеблеохватывающие. Среди крестоцветных встречаются растения как совершенно
голые, так и опушенные простыми или вильчато или звездчато разветвленными
волосками. Многолучевые звездчатые волоски зачастую напоминают чешуйки. В
опушении участвуют также железистые волоски и так называемые мальпигиевые
волоски - распростертые, двураздельные, прикрепляющиеся серединой. Для
крестоцветных характерны верхушечные кистевидные или щитковидные, обычно (или
за редким исключением) безлистные соцветия, которые иногда бывают сильно
укороченными, почти головчатыми, или же, наоборот, вытянутыми, колосовидными.
Необычный облик имеет американский каулантус вздутый (Caulanthus inflatus,), у
которого ось соцветия сильно веретеновидно утолщена и сидящие на ней цветки, а
затем плоды создают впечатление каулифлории. Цветки обычно лишены как
прицветников, так и прицветничков, не крупные, зачастую очень мелкие,
невзрачные, но немало также красиво расцвеченных, придающих растению большую
декоративность.
По своему строению они крайне
однообразны. Чашелистики, расположенные в два круга (по 2), у основания могут
быть мешковидными, и в таких случаях в эти вместилища стекает нектар. Лепестков
также 4, свободных, расположенных крестообразно (откуда и название крестоцветные).
В окраске лепестков преобладают желтый и белый цвета, но нередки также растения
с фиолетовыми, розоватыми, вплоть до пурпурных цветками. Лепестки в основном в
верхней части более широкие. Они в большинстве случаев цельные или выемчатые,
но есть среди крестоцветных также виды с лопастными (североамериканский род
вареа - Warea), перисторассеченными и даже реснитчато-бахромчатыми (у
мексиканской орнитокарпы - Ornithocarpa, например) лепестками. Тычинок обычно
6, расположенных в 2 круга. Из них 2 боковые (наружный круг) короткие, 4
срединные более длинные. Иногда срединные срастаются по две своими нитями. В
редких случаях все тычинки одинаковой длины или же по 3 разной длины. Число их
иногда может сокращаться до 4 и даже до 2 или же, как у долгонога
(Macropodium), достигает 10. У ряда видов тычинки снабжены придатками или же их
нити разрастаются в виде зубцов и крыльев. Гинецей из 2 плодолистиков. По шву
срастания плодолистиков образуется ложная перегородка, делящая завязь на 2
гнезда.
Обычно завязь сидячая, но у
некоторых видов она сидит на довольно длинном гинофоре (сходство с
каперсовыми). Особенности строения семязачатков играют немаловажную роль в
систематике крестоцветных. Семядоли обычно плоские, но бывают и вдоль
сложенными, как у капусты, реже поперек сложенными, как у гелиофилы
(Heliophila), или спирально закрученными (свербига - Bunias). По расположению
корешка зародыша по отношению к семядолям они бывают краекорешковыми и
спинкокорешковыми.
Если строением всех остальных
органов крестоцветные довольно однообразны, то этого нельзя сказать об их
плодах, признаки строения которых наиболее широко используются в систематике
семейства (рис. 31). Удлиненные плоды, длина которых значительно превышает
ширину, называются стручками, короткие же - стручочками. И те и другие могут
быть раскрывающимися двумя створками или нераскрывающимися. У раскрывающихся
плодов после опадания створок на плодоножках остается рамка (как у некоторых
каперсовых), перетянутая ложной перегородкой. Большой популярностью, например,
пользуются виды лунника (Lunaria), рамки крупных овальных стручочков которого
весьма декоративны. У нераскрывающихся стручочков часто створки сильно
уплотняются и стручочки становятся ореховидными. Особый интерес представляют
двучленные плоды, состоящие из верхнего, всегда нераскрывающегося членика и
нижнего раскрывающегося или нераскрывающегося. В одних случаях верхний членик
бывает бессемянным, в других нижний, в большинстве случаев оба членика содержат
семена. Среди двучленных плодов также различаются стручки или стручочки. Плоды
крестоцветных сильно варьируют также по величине, форме створок и различными
выростами на них.
Крестоцветные приспособлены как к
перекрестному опылению, так и к самоопылению. Основными опылителями являются
мухи, пчелы, шмели; некоторые виды, например левкоя (Matthiola) или вечерницы
(Hesperis,), опыляются в ночное время бабочками. Пчелы привлекаются запахом
медоносных видов, а также наиболее яркими цветками. Те виды, у которых цветки
мелкие, невзрачные, посещаются преимущественно мухами. Привлечение насекомых
достигается также цветовыми контрастами, иногда возникающими в процессе
цветения и плодоношения. Так, у некоторых видов с неприметными мелкими
цветками, например у веснянки (Erophila), мелкие белые лепестки начинающих плодоносить
нижних цветков соцветия не опадают, а вдвое увеличиваются и прижимаются к
незрелым плодам, имеющим фиолетовый оттенок. Этим создается как бы ореол вокруг
начинающих распускаться цветков. В другом случае, например у ярутки полевой
(Thlaspi arvense), у которой цветки также мелкие, белые, у отцветающих цветков
чашелистики становятся желтыми. У видов иберийки (Iberis) броскость
обеспечивается значительно более крупными наружными лепестками краевых цветков
соцветия, наподобие многих зонтичных. У некоторых видов гулявника (Sisymbrium),
бурачка (Alyssum), зубянки (Dentaria) этот эффект достигается за счет того, что
лепестки цветков с уже завязавшимися плодами не опадают, начинают увеличиваться
в размерах, тем самым привлекая насекомых к остальным распускающимся цветкам.
Перекрестное опыление у
крестоцветных обеспечивается благодаря присущей им дихогамии. Для большинства
из них характерна протогиния, протандрия наблюдается крайне редко. В тех
случаях, когда перекрестное опыление по какой-либо причине не может
осуществиться (обильные дожди, сильная жара, отсутствие опылителей),
крестоцветные опыляются благодаря способности к самоопылению (автогамия).
Механизм комбинированного опыления можно наблюдать, например, у горчицы полевой
(Sinapis arvensis) или у сердечника лугового (Cardamine pratense). В начале
цветения пыльники длинных тычинок поворачиваются кнаружи, вследствие чего их
пыльца не попадает на рыльце своего цветка, но может прилипнуть к бокам
насекомых-опылителей, проникающих в глубь цветка к основанию тычинок за
нектаром. Однако если рыльце не опылялось чужой пыльцой, то к концу цветения
его опыляют короткие тычинки, которые за это время достигают одного с ним
уровня. В ненастную же погоду, когда насекомых нет, пыльники длинных тычинок не
отворачиваются и опыляют рыльце своего цветка.
Есть среди крестоцветных и такие
растения, у которых в начале цветения тычинки целиком отклоняются кнаружи, а
затем приподнимаются, приближают пыльники к рыльцу и опыляют его. У
кресс-салата (Lepidium sativum), чесночника черешкового (Alliaria petiolata),
брайи альпийской (Braya alpina) в начале цветения все тычинки короче рыльца,
затем 4 из них удлиняются и соприкасаются пыльниками с рыльцем. Однако лишь
одна тычинка опорожняет пыльцу на свое рыльце, остальные пыльники раскрываются
позже, сохраняя пыльцу для перекрестного опыления.
Можно привести и такие примеры,
когда у одних видов одного и того же рода преобладает самоопыление, у других -
перекрестное. Так, ярутка альпийская (Thlaspi alpina) всегда способна к
самоопылению, так как к концу цветения тычинки наклоняются над рыльцем. И
наоборот, ярутка горная (T. montana) преимущественно перекрестноопыляющаяся,
так как у большинства растений тычинки короче рыльца. Исключительно
перекрестноопыляющиеся растения можно встретить у резухи Констанца (Arabis
constancii): рыльца у них выставляются из почки еще до распускания цветка и в
дальнейшем, когда тычинки достигают его уровня, оно от них отворачивается в
сторону так, что не может быть опылено их пыльцой. У таких растений вероятность
самоопыления исключается еще и биохимической несовместимостью пыльцы и
поверхности рыльца - своя пыльца не прорастает.
Среди крестоцветных есть также
сугубо самоопыляющиеся растения. К их числу относятся никогда не посещаемые
насекомыми виды австралийского рода стенопеталум (Stenopetalum), у которых
иногда даже образуются клейстогамные цветки. Это можно рассматривать как
приспособление к суровым условиям Западной и Южной Австралии, которые не всегда
благоприятствуют опылению. У другого австралийского растения - геококкуса
крошечного (Geococcus pusillus,) - все цветки клейстогамные. Благодаря длинным,
направленным вниз цветоножкам они зарываются в землю и там плодоносят
(геокарпия). Частичная клейстогамия характерна для бразильского сердечника
марьелистного (Cardamine chenopodiifolium), у которого, помимо нормальных
цветков верхушечного соцветия, у основания стебля образуются клейстогамные
цветки, также зарывающиеся в землю. В редких случаях, при чрезмерном
увлажнении, затоплении, клейстогамия проявляется у некоторых видов клоповника
(Lepidium), шильника водяного (Subularia aquatica), при повышенной сухости -
горчицы полевой.
Как совершенно исключительное
явление для крестоцветных можно рассматривать анемофилию, которая наблюдается
обычно у безлепестной кергеленской капусты, или принглеи (Pringlea
antiscorbutica,). Успешному ветроопылению этого островного субантарктического
вида способствуют длинные, выставляющиеся из цветка тычинки, длинные нитевидные
сосочки на рыльце и густое колосовидное соцветие.
К распространению плодов и семян
крестоцветные приспособлены довольно разнообразно. Многие из них относятся к
числу анемохоров. Это в основном виды с крылатыми или пузыревидно вздутыми
плодами, многие виды с мелкими легкими семенами, легко разносимыми ветром, или с
семенами, отороченными крылом. Иногда верхние членики двучленных плодов опадают
вместе с одной из створок нижнего членика или с частью перегородки, что также
повышает парусность.
Есть среди крестоцветных также целый
ряд видов, имеющих на плодах крючковидные выросты. Благодаря этому они
цепляются за шерсть животных и разносятся ими. Из зоохорных видов весьма
любопытен мирмекохорный клоповник пузырчатый (Lepidium vesicarium), растения
которого часто концентрически располагаются вокруг муравейников, что можно
видеть на Араратской равнине в Армении.
В некоторых случаях семена
разбрасываются благодаря «усилиям» самого растения. Так, у сердечника недотроги
(Cardamine impatiens) и сердечника шершавого (C. hirsuta) створки стручков
раскрываются с такой силой, что семена отлетают на значительное расстояние.
Довольно необычен другой вид сердечника, у которого, помимо стручков, в пазухах
листьев образуются бурые луковички, которые, опадая, прорастают. Широкой
известностью как перекати-поле пользуется так называемая иериохонская роза, или
анастатика (Anastatica hierochimtica). У этого небольшого однолетнего растения,
произрастающего в пустынных областях Западной Азии и севера Африки, плоды
созревают к началу засушливого сезона. К этому времени его многочисленные ветви
плотно сжимаются и округлые плоские стручочки остаются внутри комочка. Приняв
шаровидную форму, высохший стебель зачастую отрывается ветром от корня и
перекатывается. С наступлением дождей смоченные ветви вновь выпрямляются, этим
и напоминая распускающуюся розу. Именно тогда, при обильном увлажнении,
стручочки вскрываются (гигрохазия) и рассеивают семена. Гигрохазия вообще
присуща большинству крестоцветных с трудно раскрывающимися плодами. Семена же
нераскрывающихся плодов, защищенные от неблагоприятных условий плотным
футляром, прорастают лишь после его сгнивания. Для многих видов,
приспособленных к сухим условиям, характерно ослизнение семенной оболочки
(миксоспермия). К слизи прилипают мельчайшие частицы почвы, которые закрепляют
семена и предохраняют их от заноса в несвойственные условия среды.
Одной из особенностей многих
крестоцветных, значительно повышающей их приспособительные возможности,
является гетерокарпия в самых разнообразных ее проявлениях. В одних случаях
разнятся части плода (гетероартрокарпия), как это наблюдается у многих видов с
двучленными плодами, в других случаях - плоды целиком. Гетерокарпия
обеспечивает комбинированные способы распространения, а также более надежную
сохранность семян и возможность их прорастания при изменчивых условиях. Одним
из примеров комбинированных антропо-, гидро- и анемохории могут служить
особенности распространения двучленных плодов морской горчицы (Cakile
maritima), обитающей на морских побережьях Обе части плода содержат по одному
семени. Верхние членики благодаря сильно развитой губчатой ткани, снаружи
покрытой толстым кожистым слоем, хорошо держатся на воде и разносятся морскими
течениями. Нижние членики остаются на стеблях, которые после усыхания
отрываются от корня и перекатываются ветром. Поскольку морская горчица часто
растет вблизи портов, верхние части ее плодов зачастую вместе с грузом попадают
на судна и разносятся на дальние расстояния. Именно таким путем «уроженка»
Средиземноморья морская горчица в настоящее время широко распространена за
пределами Старого Света и успешно натурализовалась в Америке и Австралии, куда
проникла вместе с первыми колонистами
Этому, несомненно, способствовала и
ее высокая жизнеспособность, о чем свидетельствует один из любопытных
экспериментов природы. В ноябре 1963 г. в Атлантическом океане, в 20 милях
южнее Исландии, вследствие извержения подводного вулкана образовался новый
остров. Первым сосудистым растением на этом острове оказалась морская горчица,
обнаруженная там уже в июле 1965 г. Морскими течениями распространяются также
плоды катрана морского (Crambe maritima).
Хозяйственное значение крестоцветных
трудно переоценить. Овощные, масличные, кормовые и медоносные культуры имеют
среди них наиболее широкую известность, но основная роль принадлежит, конечно,
капусте во всем многообразии ее сортов. Капусту возделывали еще в
доисторические времена, и первые сведения о ней восходят к неолиту. Многие
исследователи, начиная с Ч. Дарвина, считают, что все существующие в настоящее
время культурные формы капусты происходят от дикорастущей формы капусты
огородной (Brassica oleracea), другие - от рассматриваемого в качестве
самостоятельного вида капусты лесной (Brassica sylveslris), третьи связывают их
с целым рядом средиземноморских видов. Ни одно растение в течение нескольких
тысячелетий не дало человеку столь обширного материала для отбора, как капуста.
Наибольшей популярностью пользуется капуста огородная, множество форм и сортов
которой возделывают на всех континентах. Из них капуста кочанная - основное
пищевое растение стран умеренных широт. Неоспоримы вкусовые качества таких
сортов, как кольраби, цветной капусты и ее разновидности брокколи.
Многие местные сорта особо
предпочитаются населением отдельных стран. Так, одними из древнейших культурных
растений, возделываемых в Китае и Японии, являются капуста китайская (B.
chinensis) и капуста пекинская (B. pekinensis). Как овощные растения среди
крестоцветных широко известны также различные сорта редьки и редиса (Raphanus
sativus), как острые приправы - хрен (Armoracia rusticana) и горчица сарептская
(Brassica juncea). Одной из возделываемых садово-огородных культур является
кресс-салат, в больших масштабах выращиваемый на Кавказе. В качестве салата
употребляют также ряд дикорастущих крестоцветных, как, например, ложечница
(Cochlearia), индау (Eruca sativa), сурепка (Barbarea vulgaris), жеруха
(Nasturtium officinale) и многие другие, а пастушью сумку (Capsella
bursa-pastoris) уже более 100 лет в Китае разводят как овощ. Молодые побеги и
черешки листьев катрана морского, или морской капусты (Crambe maritima), часто
употребляют подобно спарже, а в Средней Азии из корней катрана Кочи (C.
kotschyana) изготавливают муку, из которой выпекают лепешки. Большое
хозяйственное значение имеет ряд возделываемых масличных культур: рапс
(Brassica napus var. napus), горчица сарептская, горчица черная (Brassica
nigra), горчица белая (Sinapis alba), рыжик (Camelina saliva), катран
абиссинский (Crambe abyssinica). Из них в умеренных широтах наиболее урожайное
масличное растение - рапс, семена которого содержат до 50% масла. Оно имеет
сугубо техническое применение - его используют при закалке сталей, после
специальной обработки оно хорошо вулканизируется, образуя каучукообразную массу
(фактис), которую применяют для смягчения твердых каучуков и изготовления карандашных
резинок. Масло горчицы сарептской имеет пищевое применение, главным образом в
кондитерской и хлебопекарной промышленности и при изготовлении маргарина и
консервов, а порошок (жмых) представляет собой столовую горчицу. Рыжик -
единственное культурное растение среди крестоцветных, дающее полувысыхающее
масло. Его используют в мыловарении, для изготовления олифы и как смазочное для
тракторов. В США как жиро-масличное вводится в культуру высокоурожайная
лекерелла Фендлера (Lesquerella fendleri), семена которой не осыпаются и
поддаются уборке комбайном. Ее в засушливых районах рекомендуют даже вместо
пшеницы. Большинство масличных культур одновременно являются прекрасными
медоносами. Немало медоносных и эфиромасличных растений есть и среди дикорастущих
крестоцветных.
Такие ценные кормовые растения, как
брюква (Brassica napus var. napobrassica), репа и турнепс (Brassica rapa),
также принадлежат к крестоцветным. Кроме того, в качестве зеленых кормов
высевают кормовую капусту, рапс и пергу (гибрид рапса и кормовой капусты).
Список литературы
.Ботаника. Яковлев Г.П., Челомбитько В.А. М.: СпецЛит , 2008
.Ботаника: В 4 т. Систематика высших растений/ Под ред. Тимонина
А.К. М.: Academia, 2009.
.Зенькова Н.Н. Основы ботаники, агрономии и кормопроизводства. М.:
ИВЦ Минфина, 2009.
.ЖИЗНЬ РАСТЕНИЙГл. редактор член-корреспондент АН СССР проф. А.А.
ФедоровМ., «Просвещение»
.Жизнь растений. Альфред Брем; Эксмо 2010г.
Похожие работы на - Типы деления клетки. Мегаспорогенез
|