Дождевые черви как представители макрофауны почв
Дождевые черви как представители макрофауны почв
1.
Видовой состав и численность
Почвенно-биологическое
значение дождевых червей (Lumbricidae) было установлено в конце прошлого века Хенсеном и Дарвином. В
основательных трудах этих ученых уже рассматривались или по меньшей мере
затрагивались важнейшие вопросы влияния биологии дождевых червей как раз на
почвы сельскохозяйственных угодий. В последующем в разных странах были
проведены многочисленные исследования отдельных вопросов биологии дождевых
червей, которые расширили первоначальные сведения. Это касается прежде всего
выяснения таксономии дождевых червей, так как очень скоро было обнаружено,
насколько различны образ жизни и вместе с этим почвенно-биологическое значение
отдельных видов.
В среднеевропейской
области на сенокосах и пастбищах вряд ли встречается больше 10-15 видов
дождевых червей, а на полях - еще меньше, если не учитывать некоторые редко
встречающиеся виды. Они принадлежат либо к родам с красной окраской - Lumbricus, Dendrobaena и Eisenia, которые, за исключением L. terrestris (=L. herculeus), обитают преимущественно в подстилке и верхних, богатых гумусом
горизонтах, либо к родам Allolobophora, Octolasium и Eiseniella, имеющим окраску от серой до зеленоватой, так как красные пигменты
у них отсутствуют. У двух из этих шести родов в сельскохозяйственных ландшафтах
Центральной Европы имеется лишь по одному имеющему значение виду. Eisenia foetida - навозный червь, характерный для навозных и компостных куч,
первоначально обитал в лесной подстилке и не может существовать на
обрабатываемых землях. Eiseniella tetraedra встречается на берегах
рек и озер, он ведет почти водный образ жизни и может встречаться лишь на
переувлажненных лугах. Остается, следовательно, еще четыре рода, важнейшие виды
которых можно разделить на три группы по размеру: крупные виды - Lumbricus terrestris, Allolobophora longa (=A. terrestris) и Octolasium cfaneum; виды среднего размера - A. rubellus, A. caliginosa, O. lacteum; к мелким видам относятся
L. castaneus, A. rosea, A. chlorotlca, Dendrobaena rubida (включая подвид subrubicunda), D. octaedra.
Полностью
соответствующий спектр видов имеется и на северо-востоке США, как показали
исследования в штате Мэн. Туземные виды дождевых червей вымерли там в период
оледенения.
Распределение
различных видов по глубине в почвах сельскохозяйственных угодий легче всего
понять по их распределению в лесных почвах. В частности, там в растительных
остатках обитают L. rubellus, L. castaneus и Dendrobaena, как типичные потребители гумуса. Эти виды менее хорошо выживают в
почвах сенокосов и пастбищ, но если на пастбищах они находят достаточно
экскрементов животных, они охотно концентрируются под ними; на пашне они
встречаются еще реже или вовсе отсутствуют. Из червей, обитающих собственно в
почве, A. chlorotica и A. rosea как в лесах, так и на лугах предпочитают верхние слои почвы, A. caliginosa и оба вида Octolasium - средние слои, a A. terrestris и A. tonga могут уходить на глубину
1-2 м. Однако и О. суапеит и A. caliginosa также могут проделывать свои ходы до подпочвы. На пашне при
вспашке верхний и средний слои перемешиваются. Там в общем, кроме обоих
обитателей глубоких слоев, обитают еще только A. caliginosa и A. rosea и в очень сырых местах A. chlorotica в большем количестве. Горизонтальное распределение червей может
быть столь же неравномерным, как и у энхитреид. Оно зависит от влажности и
температуры почвы, растительного покрова и плотности популяций отдельных видов.
Если даже число
особей не очень показательно, тем более, что между видами и между особями
разного возраста в пределах одного вида имеются очень большие различия в
размерах, то все же оно позволяет составить известное представление о
количествах червей в почве. На сенокосах и пастбищах насчитывают 5-12 млн.
дождевых червей на 1 га (крайние значения - от 1 до 200 млн.). На пашне в
умеренном климате имеется в среднем 1-4 млн. червей на 1 га (причем минимум
равен 100 тыс., а максимум -19 млн.). Более удобным критерием является биомасса
червей, которая на лугах составляет 1000-4000 кг/га. В среднем она равна весу
крупного рогатого скота, пасущегося на равной площади при нагрузке пастбища 3
головы на 1 га и с суммарным весом коров 2000 кг. На пашне биомасса червей
колеблется в пределах 50-500 кг/га; однако как на лугах, так и на пашне
отмечали значительно больший вес червей.
. Размножение
Изучение числа хромосом показало
наличие во всех родах, за исключением Lumbricus, полиплоидных рас или видов, а следовательно, весьма часто
встречается облигатный партеногенез. Во всяком случае это относится к A. rosea, A. caliginosa (подвид trapezoides), О. суапеит и D. rubida (подвид subrubicunda), а также, вероятно, к О.
lacteum и D. octaedra, у которых не удалось обнаружить наполненных сперматек или
наблюдать копуляцию. У обычного L. terrestris спаривание происходит на
поверхности почвы.

Рис. 1. Кокон, отложенный дождевым
червем Eisenia foetida в оболочке из
одновременно выделенной червем слизи.
червь дождевой
биологический
Дождевые черви
откладывают яйца в кокон, образуемый из слизистых выделений кожных желез,
содержащий питательную жидкость и сбрасываемый червем с головного конца тела
(рис. 1). Обычно в каждом коконе имеется лишь одно яйцо, у Е. foetida - два или три. У красноокрашенных обитающих на поверхности почвы L. rubellus, L. castaneus, D. rubida и Е. foetida каждый червь откладывает
ежегодно по 70-100 коконов, у L. terrestris, A. caliginosa и A. chlorotica - по 30-40 коконов, а у A. rosea, A. longa и Octolasium - еще меньше. Дождевые
черви с красной пигментацией, в основном обитающие в лесных почвах,
размножаются наиболее быстро. Молодые черви покидают кокон после того, как
израсходуют всю питательную жидкость, и если почва достаточно влажна.
Длительность развития в коконе различна у разных видов и составляет у Е. foetida 2-3, у D. rubida в среднем 4 недели, а у
большинства других видов (по крайней мере в природных условиях) может длиться
даже 2-3 месяца. Развитие червя с момента выхода из кокона и до половой
зрелости, признаком которой является образование пояска, занимает в природных
условиях от полугода до года; как правило, черви ежегодно дают только одно
поколение. При благоприятных и выровненных условиях температуры и увлажнения в
лаборатории половая зрелость может наступить раньше. Продолжительность жизни не
превышает 2-3 лет и в значительной степени определяется обилием врагов. При
выведении червей в лаборатории длительность жизни у обоих крупных видов
достигала 5 лет и в одном случае даже 10 лет.
При благоприятных внешних условиях,
например, в компостных кучах размножение червей может происходить в течение
всего года, так что многие поколения червей следуют одно за другим, как у Е.
foetida. Обычно же в размножении
червей можно наблюдать периодичность, обусловленную климатом, который определяет
максимум и минимум активности размножения. Существование, по меньшей мере, у
некоторых видов (например, Allolobophora longa) чисто эндогенной
облигатной диапаузы, предполагаемой рядом авторов, сомнительно. Вероятно, также
и здесь сказываются факторы внешней среды. Насколько тесно зависит
периодичность размножения от условий внешней среды, видно из сравнения,
проведенного в Англии на лугопастбищных угодьях, где пасли или не пасли скот.
На лугах, где не пасутся животные, размножение дождевых червей, видимо как
результат влияния мягкого морского климата, происходит в течение всего года,
причем максимумы приходятся на периоды с октября по декабрь и с марта по
апрель. На используемых для пастьбы угодьях черви с поясками встречаются только
зимой и весной, но чаще всего в декабре и январе. Чем более континентальный
характер имеет климат, тем яснее выражены периоды покоя летом и зимой, когда
черви, свернувшись в клубок, впадают в анабиоз в расширениях в конце своих
ходов (рис. 2). Эти расширения находятся в почве на глубине 20-50 см, а у
крупных видов еще глубже; они покрыты изнутри слизистыми выделениями кожных
желез и копролитами червей, вход в эти камеры часто закрывается копролитами
червей. Благодаря этому потеря воды минимальна.
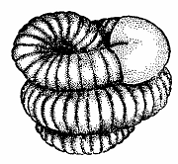
Рис. 2. Дождевой червь в состоянии
летней или зимней диапаузы
Также и в
тропическом муссонном климате Бирмы и во влажном субтропическом климате
северо-востока Индии дождевые черви (из совершенно иных родов) активны только в
дождливые периоды: в Рангуне - с мая по октябрь, в Аллахабаде - с июля до
сентября. Жаркое и холодное время года черви переживают в неактивном состоянии.
Исключение представляют только черви в сырых местообитаниях.
.
Почвенно-биологическое значение
Специфический образ
жизни дождевых червей, которые пронизывают почву сетью своих ходов, откладывают
часть перемешанных с почвой экскрементов на поверхности почвы и активно
участвуют в образовании гумуса, несомненно имеют важное значение для повышения
плодородия почвы, которое нельзя недооценивать - Благодаря деятельности червей
корни растений проникают в подпочву, органические материалы в почве быстрее
разлагаются и распределяются более равномерно. Повторно следует подчеркнуть,
что лишь формы, обитающие в минеральных почвах, способствуют перемешиванию
почвы и ее аэрации. Виды, обитающие в перегнойных почвах, реже встречающиеся на
пашне, имеют большее значение для химической переработки органического опада.
В пахотных почвах с
оптимальными условиями (травы, культуры на зеленое удобрение) количество ходов
дождевых червей достигает 1000 на 1 м2, а на полях с более частой
обработкой только 100-300 ходов на 1 м2, причем следует отметить,
что каждый червь проделывает много ходов. Такая система ходов в верхних слоях
почвы состоит преимущественно из горизонтальных каналов, но крупные виды
проделывают и вертикальные ходы до глубины более 1,5 м, идущие, следовательно,
глубоко в подпочву. Эти каналы важны для лучшей циркуляции воздуха и воды в
почве. Внутренние поверхности ходов покрыты и укреплены выделениями червей. Это
копрогенное покрытие стенок пронизывается густой сетью корней. Именно
увеличение скважности подпочвы благодаря ходам дождевых червей важно для
хорошего роста многих культурных растений. При этом меньшее значение имеет
общий объем пор, чем их распределение и взаиморасположение пустот в почве.
Подпочва, пронизанная ходами, отличается от незаселенного почвенными животными
исходного материала не большим объемом пор, а иным их расположением, так как
черви только перемещают твердые частицы. Живые корни растений в подпочве можно
найти лишь в ходах, оставшихся от прежней растительности или проделанных
дождевыми червями (рис. 3). На поле пшеницы еще на глубине 1,2 м было
обнаружено по 12-14 ходов червей диаметром 0,5 см на 1 дм2.

Рис. 3. Профиль почвы пастбища с
ходами Lumbricus terrestrls. Корневища пырея
прорастают вниз по брошенным ходам.
Дождевые черви питаются отмершими
частями растений или богатой перегноем почвой, содержащей детрит, бактерии,
водоросли, грибы и их споры, простейших и нематод. Микрофауна полностью
переваривается и поэтому, в противоположность микрофлоре, отсутствует в
копролитах червей. В их кишечнике происходит механическое и физиологическое
преобразование пищи, и у некоторых видов червей установлено даже наличие
целлюлозы. Размер частиц в комочках копролитов большей частью соответствует
размерам частиц окружающего мелкозема, однако в отдельных случаях в глотке и
мускульном желудке может происходить дальнейшее измельчение частиц почвы.
Имеющиеся в складках стенок глотки известковые железы выделяют конкреции углекислого
кальция в кипечник и оказывают влияние, таким образом, не только на равновесие
кислот и оснований в крови, но и на рН копролитов червей. Копролиты всегда
бывают менее кислыми или щелочными, чем окружающая почва, и их реакция, обычно
сдвигается в нейтральную сторону. Копролиты червей обладают большей
водопрочностью, чем обычные почвенные комки, и, в частности, копролиты червей
из луговых почв более прочны, чем червей на пашне; копролиты червей из
гумусированных верхних слоев почвы прочнее, чем подпочвы, которая содержит лишь
немного органического вещества. Частички почвы в кишечнике цементируются
большей частью слизью микроорганизмов, поглощаемых червями вместе с пищей.
Копролиты червей содержат больше углерода и общего азота, чем первичные почвенные
комки, образующиеся при механическом воздействии (выветривание, обработка
почвы, деятельность кротов), однако отношение C/N у них шире, потому что они больше
обогащаются углеродом, чем азотом. Черви A. caliginosa выделяют 6% поглощенного азота уже в доступной растениям форме.
Хотя часто бывает,
что копролиты дождевых червей содержат больше бактерий, чем обычная почва
(иногда на 33% больше), но исследования определенных питательных веществ в
почве показали, что численность микроорганизмов зависит от типа пищи. Почвы с
легкопереваримым материалом (глюкоза) после прохода через кишечник Е. foetida содержат даже меньше микроорганизмов. Напротив, в копролитах
червей из почвы, содержащей труднодоступные растительные остатки, численность
микроорганизмов возрастала. Внесенная в почву Escherichia (= Bacterium) coli не выживала, но другие микроорганизмы, прежде всего актиномицеты,
сильно размножались, конечно в зависимости от характера имеющихся питательных
веществ. Стимуляция развития части микрофлоры объясняется обеспечением
микроорганизмов пищей, благодаря производимому червями перемешиванию и переводу
в доступную форму органических материалов. Другими авторами также отмечалась
стимуляция развития актиномицетов, образующих прочные почвенные комки, и
подавление аэробных спорообразующих бактерий (например, Bacillus subtilis) в кишечнике червей. Вероятно, некоторые бактерии подавляются
действием антибиотиков, выделяемых определенными актиномицетами. Прохождение
пищи через кишечник длится 1-2 дня, т.е. она находится там достаточно долго для
развития подобных процессов. Кроме того, в кишечнике дождевых червей происходит
ускоренное разложение ингибиторных веществ из растительных остатков. Такие
вещества, замедляющие прорастание семян, были выделены из соломы и стерни зерновых.
Наконец копролиты червей удерживают больше воды, чем окружающая почва: так, на
суглинистых почвах воды в них было на 46% больше, а на песчаных почвах - на
114% больше, чем в почве. В копролитах червей, в зависимости от характера пищи,
со временем сильно развиваются не только актиномицеты, но также и другие
почвенные грибы. Мицелий этих грибов значительно повышает прочность почвенных
комков.
О количествах потребляемой червями
пищи дают представление следующие результаты. Во влажном суглинке в термостате
при 15 °С A. caliginosa в Австралии потребляли
200-300 мг сухой массы почвы в сутки на 1 г собственного веса. Прохождение пищи
через кишечник занимало 20 час. При наличии выбора черви потребляют 10 мг сухой
массы навоза в сутки на 1 г веса тела вместе с небольшим количеством листьев.
При таком питании их вес увеличивается быстрее, чем при питании одними частично
разложившимися листьями, так как микроорганизмы, изобилующие в навозе, либо
повышают доступность пищи, либо перевариваются вместе с ней. Благодаря
тщательному перемешиванию минеральных частиц и органического вещества в
кишечнике многих видов дождевых червей образуются устойчивые
глинисто-перегнойные комплексы, которые без разрушения веществ не могут быть
полностью разделены ни химическими, ни физическими средствами. Эти комплексы
представляют собой важную составную часть мулля - наиболее ценной формы гумуса.
Такими же создателями гумуса, помимо дождевых червей, являются энхитреиды, а в
тропических странах - термиты, тогда как в кишечнике большинства других
почвенных сапрофагов образуется модер, в котором не полностью разложившиеся
органические и неорганические компоненты можно еще разделить механическим
путем.
Литература
1. Тишлер В. Сельскохозяйственная экология. М., «Колос», 1971.
. Основы общей и сельскохозяйственной экологии: Ю.А. Захваткин -
Санкт-Петербург, Мир, 2003 г. - 360 с.
. Лекции по экологии: О.В. Богданкевич - Москва, ФИЗМАТЛИТ, 2002
г. - 208 с.
. Общая экология: М.В. Гальперин - Санкт-Петербург, Форум,
Инфра-М, 2007 г. - 336 с.