Структура и функции белков
Содержание
1.
Дайте характеристику физических методов исследования строения белков. Приведите
примеры
.
Напишите химическую формулу пептида серил-лейцил-треонил-пролина
.
Приведите примеры зависимости биологической активности белков от их первичной
структуры
.
Напишите химические формулы уридиловой и псевдоуридиловой кислот. Укажите, в
состав каких нуклеиновых кислот они входят. Объясните роль псевдоуридиловой
кислоты как структурного мономера
.
Напишите уравнения реакций, в которых участвуют качестве коферментов тиаминый
рофосфат, липоат, пиридоксальфосфат
.
Напишите начальный этап ферментативного фосфоролиза аланиновой т-РНК,
выделенной из пекарских дрожжей. Назовите фермент, катализирующий процесс
фосфоролиза и образовавшиеся продукты реакции
.
Составьте уравнение реакции переаминирования гистидина и глиоксиловой кислоты.
Покажите на данных примерах механизм действия пиридоксальфермента
.
Используя схему превращений метаболитов в цикле три-и дикарбоновых кислот,
найдите этапы дегидрирования метаболитов. Напишите химические уравнения реакций
и, учитывая особенности передачи энергии на синтез АТФ через систему
дыхательных ферментов подсчитайте число синтезированных молекул АТФ за один
цикл превращения
.
В биосинтезе пальмитиновой кислоты одной из промежуточных стадий является
превращение: капронил-S-КОА=каприл-S-КОА. Напишите уравнение реакции и укажите
ферменты ускоряющие эти процессы
.
Охарактеризуйте окислительное фосфорилирование на примере окислительного
декарбоксилирования пировиноградной кислоты. Чем отличается окислительное
фосфорилирование от фотосинтетического?
.
Приведите примеры биологически активных производных гормона адреналина.
Напишите уравнение реакции их биосинтеза, покажите их физиологическое действие
.
Опишите уровни регуляции биосинтеза белка
Используемая
литература
белок
кислота гормон биосинтез
1. Дайте характеристику физических
методов исследования строения белков. Приведите примеры
Классификация методов не может быть абсолютно
строгой, поскольку не всегда удается выделить специфические свойства,
определяемые данным методом. Но в целом возможно оценить наиболее важные
характеристики методов исследования.
Спектроскопические методы
В большинстве этих методов измеряют зависимость
интенсивности излучения I, прошедшего через вещество или рассеянное веществом,
от частоты n, то есть определяют функцию I(n). В настоящее время диапазон
частот распространяется на значения от минимальных n ї 106 Гц в ядерном
магнитном резонансе (ЯМР) до 1019 Гц (гамма-излучение). Частоты n и длины волн
l излучения связаны простым соотношением с = ln, с - скорость света в вакууме.
Для указанного диапазона частот длины волн изменяются от l - 200 м до 10 - 14
м. Столь значительный диапазон частот (длин волн) требует различных источников
излучения и выявляет различные физические свойства вещества. Наблюдаемые
частоты соответствуют разностям энергий Е двух состояний молекул:
, 2 = (E2 - E1)/ h,
где h - постоянная Планка. Переходами между
уровнями энергии E1 и E2 «управляют» правила отбора. Это означает, что не все
переходы возможны. Из таблицы видно, что разница, например, между двумя
уровнями энергии валентных электронов (УФ-спектры - ультрафиолетовые спектры)
DЕ (УФ) = E2 - E1 значительно больше, чем DЕ (ЯМР).
Для химии важны не только абсолютные разницы DЕ,
но их изменения в различных соединениях, вызванных изменениями в составе или
под влиянием ближайшего окружения.
Наибольшее распространение для идентификации
веществ получили колебательные и электронные спектры, а также спектры ядерного
магнитного резонанса.
В колебательной спектроскопии важно иметь полный
интервал частот от очень низких (порядка 10 см - 1), характерных для крутильных
колебаний, до высоких значений (порядка 5000 см - 1). Частоты колебательных
спектров используются также для расчета силовых полей молекул, то есть для
определения различного типа сил взаимодействия атомов в молекуле. Так
называемые силовые постоянные для значительного числа групп атомов обладают
свойством переносимости, то есть постоянством в рядах сходных по строению
молекул.
Электронная спектроскопия является очень
чувствительным и удобным методом для определения спектров поглощения,
пропускания и отражения, изучения кинетики реакции, сопровождающихся
спектральными изменениями. В обычных условиях спектры имеют диффузный характер,
что ограничивает их применение веществами, имеющими хромофорные группы
(ароматические циклы, кратные связи и т.п.). Эти спектры позволяют
устанавливать наличие тех или иных групп в молекуле, то есть осуществлять
групповой анализ, изучать влияние заместителей на электронные спектры и
строение молекул, исследовать таутомерию и другие превращения.
Метод ядерного магнитного резонанса (ЯМР)
основан на взаимодействии внешнего магнитного поля с ядрами, имеющими магнитный
момент, такими как 1H, 13C, 15N, 19F, 29Si, 31P, для которых спиновое квантовое
число равно 1/2, а также для ряда ядер со спиновым квантовым числом, большим
1/2. Одни и те же ядра атомов в различных окружениях в молекуле показывают
различные сигналы ЯМР. Отличие такого сигнала ЯМР от сигнала стандартного вещества,
например тетраметилсилана, позволяет определить так называемый химический
сдвиг, который обусловлен химическим строением изучаемого вещества. В методиках
ЯМР есть много возможностей определять химическое строение веществ, конформации
молекул, эффекты взаимного влияния, внутримолекулярные превращения и т.п.
Дифракционные методы
В дифракционных методах используются волновые
свойства излучения и потока частиц электронов и нейтронов. Волновые свойства
рентгеновских лучей были открыты в 1912 году немецким физиком Лауэ. Он заложил
основы рентгеноструктурного анализа. Гипотеза о волновых свойствах частиц
выдвинута французским физиком Луи де Бройлем в 1924 году. Эта гипотеза выражена
простым соотношением между длиной волны l, массой m и скоростью u движущейся
частицы:
В 1927 году экспериментально подтверждено
явление дифракции электронов. Позднее открыта дифракция нейтронов.
В дифракционных методах измеряют зависимость
интенсивности рассеянного излучения от угла рассеяния q, то есть функцию I(q).
При этом длина волны после рассеяния не изменяется. Имеет место так называемое
упругое рассеяние.
Рентгеновские лучи, получаемые в рентгеновских
трубках, имеют длины волн порядка 0,07 - 0,2 нм. Длины волн электронных пучков
составляют величины порядка 0,005 нм. В нейтронографии потоки нейтронов
характеризуются длинами волн порядка 0,15 нм. Существенным ограничением
использования нейтронов является то, что их источник связан с ядерным
реактором.
Хотя эти три типа падающего излучения
удовлетворяют основному соотношению дифракции, они используются несколько
по-разному. Последнее объясняется различным характером взаимодействий
рентгеновских лучей, электронов и нейтронов с веществом. Наиболее сильно
рассеиваются электроны. Слабее всего рассеиваются нейтроны.
Поэтому рентгенография и нейтронография
используются для исследований кристаллов и другой конденсированной фазы в
макроскопических размерах. Электронография применяется для изучения тонких
пленок, поверхностей и газов.
Наиболее широкое применение в химии нашли два
метода:
. Рентгеноструктурный анализ, который позволяет
определять координаты атомов в трехмерном пространстве кристаллических веществ
от простейших соединений типа NaCl до сложных белков.
. Газовая электронография, с помощью которой
определяют геометрию свободных молекул в газах, то есть молекул, не
подверженных влиянию соседних молекул, как это имеет место в кристаллах.
Сопоставление данных обоих методов для одних и
тех же веществ дает возможность оценить влияние кристаллического поля на
молекулу.
Оптические методы
Оптическими методами изучают распространение,
рассеяние и поглощение света в веществе. Физические величины, которые измеряют,
представляют следующий ряд:
) n - показатель преломления: n = c/u, где с -
скорость света в вакууме, u - скорость света в веществе;
) a - угол поворота плоскости поляризации
линейно поляризованного света при прохождении через оптически активное
вещество, которое вращает плоскость поляризации падающего линейно
поляризованного света;
) r - коэффициент деполяризации, то есть отношение
интенсивности рассеянного под углом 900 света с поляризацией, перпендикулярной
плоскости падающего линейно поляризованного света I^ , к интенсивности
рассеянного света с параллельной поляризацией I|| , то есть r = I^ / I|| .
) - эффект Керра, где n|| и n^ - показатели
преломления для линейно поляризованных лучей, распространяющихся вдоль
электрического поля E|| и перпендикулярно этому полю соответственно;
) a(В ) - эффект Фарадея, где a(В ) -
зависимость угла поворота плоскости поляризации света от величины магнитного
поля В ;
) e(l) - молярный коэффициент поглощения света
как функция l и др.; этот параметр определяется также в спектроскопических
методах.
Результаты оптических методов используются для
идентификации веществ, выявления взаимного влияния атомов в молекуле, расчета
поляризуемости молекул, отнесения частот в колебательном анализе, изучения
влияния растворителя на исследуемую систему и т.д.
Масс-спектрометрия и спектроскопия электронов
Эта группа методов отличается от предыдущих тем,
что в результате взаимодействия какого-либо падающего излучения или потока
частиц на вещество измеряются потоки других частиц.
Методом масс-спектрометрии определяют
молекулярные массы, идентифицируют вещества, устанавливают химическое строение
веществ, изучают теплоты испарения и реакций, механизмы химических реакций,
измеряют потенциалы ионизации и энергии разрыва химических связей.
В методах рентгеновской электронной
спектроскопии (РЭС) и оптической электронной спектроскопии (фотоэлектронной
спектроскопии, ФЭС) падающим излучением I0 является рентгеновское или
ультрафиолетовое излучение. Однако в отличие от масс-спектрометрии измеряют
энергии потока электронов, вырванных из молекулы или вещества, то есть измеряют
I(Eэл).
Рентгеновское излучение вырывает электроны из
внутренних оболочек атомов веществ. Поэтому метод РЭС позволяет определять
энергию связи внутренних электронов остовов атомов в молекуле и веществе.
Методом ФЭС определяют последовательные потенциалы ионизации из валентной
оболочки атомов в молекуле. Оба эти метода дают возможность идентифицировать
вещества и изучать закономерности влияния атомов ближайшего окружения на
энергию связи электронов в атомах на разных орбиталях.
Диэлькометрия и магнетохимия
В зависимости от величин электрических дипольных
моментов или магнитных характеристик веществ внешние электрическое и
соответственно магнитное поля изменяют поведение вещества в этих полях по
сравнению с поведением в отсутствие поля.
Измерения диэлектрической проницаемости e
позволяют определить величину электрического дипольного момента m, который
характеризует полярность молекул. Кроме того, величина m является источником
структурной информации при использовании аддитивных расчетных схем.
Магнетохимические исследования дают возможность
оценивать по степени парамагнетизма число неспаренных электронов в атомах
вещества (парамагнитные вещества втягиваются в магнитное поле). Диамагнетики
выталкиваются магнитным полем и степень этого выталкивания обусловлена
электронным строением молекул и вещества. Особенно показательно различие
молярных диамагнитных восприимчивостей cd , параллельных и перпендикулярных
плоскости молекул бензола, нафталина и других ароматических углеводородов. Это
доказывает существование электронных токов в плоскостях ароматических молекул.
2. Напишите химическую формулу
пептида серил-лейцил-треонил-пролина
H2N-CH-CO-NH-CH-CO-NH-CH-COOH | | |
CH3 CH CH2 | | (CH3)2 C2H5
3. Приведите примеры зависимости
биологической активности белков от их первичной структуры
Белки - это высокомолекулярные соединения
(полимеры), состоящие из аминокислот - мономерных звеньев, соединенных между
собой пептидными связями. Все 20 аминокислот, встречающиеся в белках, это a
-аминокислоты, общим признаком которых является наличие аминогруппы - NН2 и
карбоксильной группы - СООН у a -углеродного атома. a -аминокислоты отличаются
друг от друга структурой группы R и, следовательно, свойствами. Все
аминокислоты можно сгруппировать на основе полярности R-групп, т.е. их
способности взаимодействовать с водой при биологических значениях рН.
Пептидные связи образуются при взаимодействии a
-аминогруппы одной аминокислоты с a -карбоксильной группой другой аминокислоты:
Пептидная связь - это амидная ковалентная связь, соединяющая аминокислоты в
цепочку. Следовательно, пептиды - это цепочки аминокислот.
Изображение последовательности аминокислот в
цепи начинается с N-концевой аминокислоты. С неё же начинается нумерация
аминокислотных остатков. В полипептидной цепи многократно повторяется группа:
-NH-CH-CO-. Эта группа формирует пептидный остов. Следовательно, полипептидная
цепь состоит из остова (скелета), имеющего регулярную, повторяющуюся структуру,
и отдельных боковых цепей R-групп. Первичная структура характеризуется порядком
(последовательностью) чередования аминокислот в полипептидной цепи. Даже
одинаковые по длине и аминокислотному составу пептиды могут быть разными
веществами потому, что последовательность аминокислот в цепи у них разная.
Последовательность аминокислот в белке уникальна и детерминируется генами. Даже
небольшие изменения первичной структуры могут серьезно изменять свойства белка.
Было бы неправильно заключить, что каждый аминокислотный остаток в белке
необходим для сохранения нормальной структуры и функции белка.
Функциональные свойства белков определяются их
конформацией, т.е. расположением полипептидной цепи в пространстве.
Уникальность конформации для каждого белка определяется его первичной
структурой. В белках различают два уровня конформации пептидной цепи -
вторичную и третичную структуру. Вторичная структура белков обусловлена
способностью групп пептидной связи к водородным взаимодействиям: C=O....HN.
Пептид стремится принять конформацию с максимумом водородных связей. Однако
возможность их образования ограничивается тем, что пептидная связь имеет
частично двойной характер, поэтому вращение вокруг нее затруднено. Пептидная
цепь приобретает не произвольную, а строго определенную конформацию,
фиксируемую водородными связями. Известны несколько способов укладки
полипептидной цепи: a -спираль - образуется внутрицепочечными водородными
связями между NH-группой одного остатка аминокислоты и CO-группой четвертого от
нее остатка; b -структура (складчатый лист) - образуется межцепочечными
водородными связями или связями между участками одной полипептидной цепи
изогнутой в обратном направлении; беспорядочный клубок - это участки, не
имеющие правильной, периодической пространственной организации. Но конформация
этих участков также строго обусловлена аминокислотной последовательностью.
Содержание a -спиралей и b -структур в разных белках различно: у фибриллярных
белков - только a -спираль или только b -складчатый лист; а у глобулярных
белков - отдельные фрагменты полипептидной цепи: либо a -спираль, либо b
-складчатый лист, либо беспорядочный клубок. Третичная структура глобулярных
белков представляет ориентацию в пространстве полипептидной цепи, содержащей a
-спирали, b -структуры и участки без периодической структуры (беспорядочный
клубок). Дополнительное складывание скрученной полипептидной цепи образует
компактную структуру. Это происходит, прежде всего, в результате взаимодействия
между боковыми цепями аминокислотных остатков.
4. Напишите химические формулы
уридиловой и псевдоуридиловой кислот. Укажите, в состав каких нуклеиновых
кислот они входят. Объясните роль псевдоуридиловой кислоты как структурного
мономера
Формула уридиловой кислоты:
C=O 9N CH 9¦ 0 9¦ 9О=С СН Н 42 0РО 43 0- О - СН
42 0 N ¦ О ¦ С 4Н 0 4Н 0 С 4Н 0 С 4-- 0 С 4 Н ¦ ¦ ОН ОН
Формула псевдоуридиловой кислоты:
C=O 9N CH 9¦ 9О=С СН Н 42 0РО 43 0- О - СН 42 0
N ¦ О ¦ С 4Н 0 4Н 0 С 4Н 0 С 4-- 0 С 4 Н ¦ ¦ ОН ОН
Данные кислоты входят в состав АТФ, ГТФ, НАД+ ,
НАДФ+ , ФАД, ФМН
Псевдоуридиловая кислота, как структурный
мономер нуклеиновых кислот выполняет роль акумулятора и переносчика энергии,
мессенджера или второго вестника в реализации клеткой внеклеточного
регуляторного сигнала, а также участвует во многих метаболических процессах в
клетке.
5. Напишите уравнения реакций, в
которых участвуют качестве коферментов тиаминый рофосфат, липоат,
пиридоксальфосфат
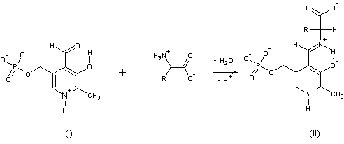
Рис
Тиаминый рофосфат в качестве кофермента
используется в реакции трансаминирования L-аланина, задействованную для
обеспечения транспорта аммиака в организме.
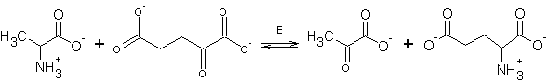
Липоат в качестве кофермента может
использоваться при замещении остатка лизина субстратом
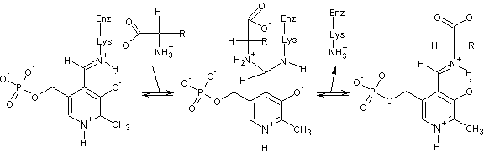
Рис
6. Напишите начальный этап
ферментативного фосфоролиза аланиновой т-РНК, выделенной из пекарских дрожжей.
Назовите фермент, катализирующий процесс фосфоролиза и образовавшиеся продукты
реакции
Рис
Процесс фосфоролиза катализируется ферментом
теломераза. В результате фосфоролиза аланиновой т-РНК, выделенной из пекарских
дрожжей образовываются тирозин специфическая РНК.
7. Составьте уравнение реакции
переаминирования гистидина и глиоксиловой кислоты. Покажите на данных примерах
механизм действия пиридоксальфермента
Реакция переаминирования гистидина и
глиоксиловой кислоты
СН2 ОН СН2О Р
СО СО
фосфорибулокиназа
Н С ОН + АТФ Н С ОН + АДФ
Н С ОН Н С ОН
СН2О Р СН2О Р
. Используя схему превращений
метаболитов в цикле три-и дикарбоновых кислот, найдите этапы дегидрирования
метаболитов. Напишите химические уравнения реакций и, учитывая особенности
передачи энергии на синтез АТФ через систему дыхательных ферментов подсчитайте
число синтезированных молекул АТФ за один цикл превращения
Превращения веществ в клетке (обмен веществ, или
метаболизм), в результате которых из сравнительно простых предшественников,
например глюкозы, жирных кислот с длинной цепью или ароматических соединений,
образуется новое клеточное вещество, можно ради простоты подразделить на три
основные группы. Сначала питательные вещества расщепляются на небольшие
фрагменты (распад, или катаболизм), а затем в ходе реакций промежуточного
обмена, или амфиболизма, они превращаются в ряд органических кислот и фосфорных
эфиров. Эти два пути переходят незаметно один в другой. Многообразные
низкомолекулярные соединения - это тот субстрат, из которого синтезируются
основные строительные блоки клетки. «Строительными блоками» мы называем
аминокислоты, пуриновые и пиримидиновые основания, фосфорилированные сахара,
органические кислоты и другие метаболиты - конечные продукты цепей биосинтеза,
иногда длинных. Из них строятся полимерные макромолекулы (нуклеиновые кислоты,
белки, резервные вещества, компоненты клеточной стенки и т.п.), из которых
состоит клетка. Эти два этапа биосинтеза клеточных веществ - синтез строи
тельных блоков и синтез полимеров - составляют синтетическую ветвь метаболизма,
или анаболизм
СН2О R СН2О
Р
фосфоглицераткиназа
СН ОН + АТФ СН ОН + АДФ
СООН СОО ~ Р
- фосфоглицериновая кислота
,3 - дифосфоглицериновая кислота
9. В биосинтезе пальмитиновой
кислоты одной из промежуточных стадий является превращение: капронил-S-КОА=каприл-S-КОА.
Напишите уравнение реакции и укажите ферменты ускоряющие эти процессы
СНСОО СН2СООН + НS~КоА
+ Н2О цитратсинтаза С(ОН)СООН + СН3СО~SКоА
С(ОН)СООН СН2СООН
лимонная кислота
Фермент, ускоряющий процесс называется
цитратсинтаза.
10. Охарактеризуйте окислительное
фосфорилирование на примере окислительного декарбоксилирования пировиноградной
кислоты. Чем отличается окислительное фосфорилирование от фотосинтетического?
Окислительное фосфорилирование - один из
важнейших компонентов клеточного дыхания, приводящего к получению энергии в виде
АТФ. Субстратами окислительного фосфорилирования служат органические соединения
- белки, жиры и углеводы.
Однако чаще всего в качестве
субстрата используются углеводы
<http://ru.wikipedia.org/wiki/%D0%A3%D0%B3%D0%BB%D0%B5%D0%B2%D0%BE%D0%B4%D1%8B>.
Так, клетки головного мозга
<http://ru.wikipedia.org/wiki/%D0%93%D0%BE%D0%BB%D0%BE%D0%B2%D0%BD%D0%BE%D0%B9_%D0%BC%D0%BE%D0%B7%D0%B3>
не способны использовать для дыхания никакой другой субстрат, кроме углеводов.
Предварительно сложные углеводы
расщепляются до простых, вплоть до образования глюкозы. Глюкоза является
универсальным субстратом в процессе клеточного дыхания. Окисление глюкозы
подразделяется на 3 этапа:
гликолиз;
окислительное декарбоксилирование
или цикл Кребса;
окислительное фосфорилирование.
При этом гликолиз является общей
фазой для аэробного и анаэробного дыхания.
Окислительное декарбоксилирование
пировиноградной кислоты:
СН3―СО―СООН →
2 СН3―СО~КоА + 2
СО2 + 2 НАД×Н2
В дыхательную цепь поступает: 10
НАДН2, каждый из которых окислясь до воды выделяет энергии достаточной для
образования 3АТФ, следовательно всего получается 30 АТФ; 2 ФАДН2 окисляясь в
дыхательной цепи каждый дает 2 АТФ, следовательно всего 4 АТФ. Кроме того, в
гликолизе и ЦТК образуется по 2 АТФ в результате субстратного фосфорилирования.
Таким образом, при полном окислении глюкозы синтезируется 38 молекул АТФ.
Характерная особенность анаэробной
фазы пентозофосфатного цикла - переход от продуктов гликолиза к образованию
фосфопентоз, необходимых для синтеза нуклеотидов и нуклеиновых кислот, и
наоборот, использование продуктов пентозного пути для перехода к гликолизу.
Важнейшим соединением, обеспечивающим такой двухсторонний переход, является
эритрозо-4-фосфат, которое является предшественником в биосинтезе ароматических
аминокислот у автотрофных организмов. Петозофосфатный цикл не является основным
путем обмена глюкозы и обычно не используется клеткой для получения энергии.
Биологическое значение
пентозофосфатного цикла заключается в снабжении клетки восстановленным НАДФ×Н,
необходимым для биосинтеза жирных кислот, холестерина, стероидных гормонов,
пуринов и др. важнейших соединений.
Ферменты пентозофосфатного цикла
используются в темновой фазе фотосинтеза при образовании глюкозы из углекислого
газа в цикле Кальвина. Пентозофосфатный путь широко представлен в природе и
обнаружен у животных, растений и микроорганизмов.
11. Приведите примеры биологически
активных производных гормона адреналина. Напишите уравнение реакции их
биосинтеза, покажите их физиологическое действие
Гормоны - это биологически
высокоактивные вещества, синтезирующиеся и выделяющиеся во внутреннюю среду
организма эндокринными железами, или железами внутренней секреции, и
оказывающие регулирующее влияние на функции удаленных от места их секреции
органов и систем организма. Эндокринная железа - это анатомическое образование,
лишенное выводных протоков, единственной или основной функцией которого
является внутренняя секреция гормонов. К эндокринным железам относятся гипофиз,
эпифиз, щитовидная железа, надпочечники (мозговое и корковое вещество),
паращитовидные железы.
Адреналин - гормон мозгового слоя надпочечников
животных и человека. Поступая в кровь, адреналин повышает потребление кислорода
и артериальное давление, содержание сахара в крови, стимулирует обмен веществ.
При эмоциональных переживаниях, усиленной мышечной работе содержание адреналина
в крови увеличивается.
Адренокортикотропный гормон (АКТГ,
кортикотропин) - гормон животных и человека, вырабатываемый гипофизом. АКТГ
стимулирует рост коры надпочечников и образование в ней гормонов -
кортикостероидов. При мобилизации защитных сил организма синтез АКТГ
усиливается.
Альдостерон - гормон животных и человека,
вырабатываемый в коре надпочечников (кортикостероид). Он регулирует минеральный
обмен в организме: стимулирует задержку ионов натрия (Na+) в крови и выведение
ионов калия (K+) и водорода (H+).
Вазопрессин (антидиуретический гормон) -
нейрогормон животных и человека, который вырабатывается в гипоталамусе,
поступает в гипофиз, а затем выделяется в кровь. Вазопрессин стимулирует
обратное всасывание воды в почечных канальцах и таким образом уменьшает
количество выделяющейся мочи (антидиуретический эффект). При недостатке
вазопрессина резко повышается выделение мочи, что может привести к несахарному
диабету. Таким образом, вазопрессин - один из факторов, определяющих
относительное постоянство водно-солевого обмена в организме. Вазопрессин
вызывает также сужение сосудов и повышение кровяного давления.
Биосинтез производных адреналина протекает по
следующим химическим реакциям:
Глюкоза + АТФ гексокиназа сахарозо-6-фосфат;
глюкозо-6-фосфат фосфоглюкомутаза
глюкозо-1-фосфат;
глюкозо-1-фосфат + УТФ уридилтрансфераза
УДФ-глюкоза + Н4Р2О7
УДФ-глюкоза УДФ-глюкоза -4- эпимераза УДФ -
галактоза;
УДФ-галактоза + глюкоза лактозо-УДФ-гликозил-
лактоза + УДФ.
12. Опишите уровни регуляции
биосинтеза белка
Регуляция биосинтеза белка - принципиальный
атрибут любой живой клетки. Регуляция необходима для поддержания баланса
разнообразных белков в клетке или организме, для изменения этого баланса в
меняющихся условиях окружающей или внутриорганизменной среды, для обеспечения
смены белков в процессах клеточной дифференцировки и развития организма, для
адекватного ответа на специфические внешние сигналы или неблагоприятные
воздействия. Синтез белков в клетке регулируется на трех уровнях: 1) путем
изменения активности генов, то есть через тотальную или избирательную модуляцию
продукции мРНК на матрице ДНК (уровень транскрипции); 2) путем изменения
активности мРНК в ее трансляции рибосомами (уровень трансляции); 3) путем
деградации мРНК посредством ее тотального или избирательного расщепления
рибонуклеазами.
Существуют три основных способа, как
регулировать трансляцию. Первый способ - позитивная регуляция на основе
сродства мРНК к инициирующей рибосоме и факторам инициации (дискриминация
мРНК). Второй способ - негативная регуляция с помощью белков-репрессоров,
которые, связываясь с мРНК, блокируют инициацию (трансляционная репрессия).
Этими двумя способами регулируются индивидуальные мРНК, то есть трансляция
каждой мРНК может специфически контролироваться независимо от других мРНК
клетки. Третий способ - тотальная регуляция трансляции всей совокупности мРНК
клетки посредством модификации факторов инициации.
Используемая литература
1. Березов
Т. Т., Коровкин Б. Ф. Биологическая химия. - М.: Высшая школа, 1990. - 208 с.
2. Кнорре
Д.Г., Мызина С.Д Биологическая химия. - М.: Высшая школа, 2000. - 198 с.
. Королёв
А.П., Гридина С.Б., Зинкевич Е.П. Основы биохимии. - Кемерово: Кемеровский
технологический институт пищевой промышленности.- Кемерово, 2004. - 92 с.
. Ленинджер
А. Основы биохимии. - М.: Мир, 1985. - 384 с.
. Спирин
А.С. Молекулярная биология: Структура и биосинтез нуклеиновых кислот. - М.:
Высшая школа, 1990. - 186 с.
. Фершт
Э. Структура и механизм действия ферментов. - М.: Мир, 1980. - 262 с.
. Элиот
В., Элиот Д. Биохимия и молекулярная биология. - М.: НИИ Биомед. химии РАМН,
1999. - 328 с.