Электрические импульсы в биологических клетках
Содержание
Введение
Электрический заряд
Электрическое поле. Закон Кулона.
Характеристики электрического поля
Мембранный потенциал покоя
Биофизические основы мембранного
потенциала покоя
Потенциал действия
Биофизические основы потенциала
действия
Распространение потенциала действия
Биопотенциалы
Мембранная теория биопотенциалов
Открытие потенциала действия
Биоэлектрические потенциалы у
растений
Круговые токи. Законы проведения
возбуждения по нервному волокну
Заключение
Список литературы
Введение
«Как ни чудесны законы и явления электричества,
выявляющиеся нам в мире неорганического или мертвого вещества, интерес, который
они представляют, вряд ли может сравниться с тем, что присуще той же силе в
соединении с нервной системой и жизнью».
М. Фарадей
В ходе изучения биологических клеток было давно
замечено, что о какой бы клетке ни шла речь, на мембране или оболочке,
окружающей клетку, наблюдаются электрические явления. При сокращении мышц ткани
регистрируются импульсы электрического напряжения. Отдельные мышцы легко
возбуждаются электрическим током, в результате чего наблюдаются их сокращения.
При измерениях, проводимых на кожном покрове, обнаруживаются электрические
заряды, которые создают своеобразный потенциальный рельеф. Оценивая его, можно
получить информацию о состоянии органов и систем, имеющих связь с данной
локальной зоной.
Электрический заряд
Электрический заряд, подобно массе, является
фундаментальным свойством веществ. Существует два типа зарядов, условно
обозначенные как положительный и отрицательный.
Каждое вещество имеет электрический заряд,
величина которого может быть положительной, отрицательной или быть равной нулю.
Например, электроны заряжены отрицательно, а протоны - положительно. Поскольку
каждый атом содержит один или более электронов и равное количество протонов,
общее число зарядов в макроскопическом объекте - чрезвычайно большое, но в
целом такой объект не заряжен или имеет небольшой заряд.
Заряд электрона является по абсолютной величине
самым маленьким.
Электрическое поле. Закон Кулона
Каждый заряженный объект образует в окружающем
его пространстве электрическое поле. Электрическое поле является видом материи,
посредством которой заряженные объекты взаимодействуют друг с другом. Пробный
заряд, внесённый в электрическое поле другого заряда "чувствует"
присутствие этого поля. Он будет притягиваться к заряду, создающему
электрическое поле, или отталкиваться от него.
Закон Кулона определяет электрическую силу F,
действующую между двумя точечными зарядами q1 и q2:
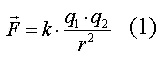
- константа, определяемая выбранными условиями;
r - расстояние между зарядами.
Согласно закону Кулона, сила действует в
направлении линии, соединяющей два заряда. Величина силы, действующей на
заряды, пропорциональна величине каждого из зарядов и обратно пропорциональна
квадрату расстояния между ними.
Электрическое поле можно представить в виде
силовых линий, показывающих направление электрических сил. Эти силы направлены
от заряда, когда он положительный, и к заряду, если он отрицательный. Если
положительный заряд поместить в электрическое поле, он подвергается действию
силы в направлении поля. Отрицательный заряд подвергается силе, направленной
противоположно направлению поля.
Характеристики электрического поля
Напряжённость электрического поля. Каждый
электрический заряд производит вокруг себя электрическое поле. Если другой
заряд q внести в это поле, то на него будет действовать сила F,
пропорциональная q и напряжённости электрического поля E:
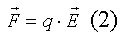
Напряжённость электрического поля (E)
в любой точке определяется как электрическая сила (F), которая действует на
положительной заряд q, помещённый в эту точку:
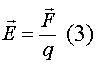
- векторная величина, то есть имеет как
величину, так и направление. Единицей измерения напряжённости является вольт на
метр [В/м].
Принцип наложения (суперпозиции) указывает, что
если электрическое поле создают множество зарядов, суммарная напряжённость
определятся сложением напряжённостей, созданных каждым зарядом, по правилам
сложения векторов.
Электрический потенциал. Чтобы переместить заряд
против действующей на него электрической силы необходимо выполнить работу. Эта
работа не зависит от пути перемещения заряда в электрическом поле, но зависит
от начального и конечного положения заряда.
Если заряд перемещается из одной точки в другую
против электрической силы, его потенциальная электростатическая энергия
увеличивается. Электрический потенциал в любой точке равен электростатической
потенциальной энергии Wp, которую имеет положительный заряд q в этой точке: φ
= Wp/q
(4).
Можно также сказать, что электрический потенциал
в точке равен работе, которую необходимо совершить против электрических сил,
чтобы переместить положительный заряд из данной точки на большое расстояние,
где потенциал электрического поля равен нулю. Электрический потенциал является
скалярной величиной и измеряется в вольтах (В).
Напряжённость электрического поля является
отрицательным градиентом электрического потенциала - показателя изменения потенциала
с расстоянием x: E→ = - dφ/dx. С
помощью приборов можно измерить разность потенциалов, но не напряжённость поля.
Последняя может быть вычислена, если использовать зависимость между E→ и Δφ
:
где Δφ
= E·l - расстояние
между двумя токами электрического поля.
Мембранный потенциал покоя
Каждая клетка превращает часть своей
метаболической энергии в электростатическую энергию. Источником электрического
поля клетки является плазматическая мембрана. Существует разность потенциалов
между внутренней и внешней поверхностями плазматической мембраны. Эта разность
потенциалов называется мембранным потенциалом.
Разность потенциалов между внутренней и внешней
средами клетки может измеряться непосредственно и довольно точно. Для этого
используют микроэлектрод, представляющий собой стеклянную микропипетку с
диаметром кончика до 1мкм, заполненную концентрированным раствором KCl.
Микроэлектрод подключают к усилителю напряжения регистрирующего устройства.
Можно измерять мембранный потенциал мышечных, нервных клеток или клеток других
тканей. Другой электрод (референтный) установлен на поверхности ткани.
Когда кончик микроэлектрода находится вне
клетки, его потенциал по отношению к референтному электроду равен нулю. Если
конец электрода погружают в клетку, прокалывая плазматическую мембрану,
разность потенциалов резко становится отрицательной. На шкале измерительного
устройства регистрируется разность потенциалов между внутренней и внешней
средами клетки. Эта разность потенциалов называется трансмембранной, или
мембранным потенциалом.
Если клетка находится в состоянии покоя, её
мембранный потенциал имеет отрицательное значение и устойчивую величину. Обычно
его называют мембранным потенциалом покоя. Мембранный потенциал покоя клеток
различных тканей составляет от - 55 милливольт (мВ) до -100 мВ
При определенных физиологических условиях могут
происходить изменения мембранного потенциала. Изменения его в положительном
направлении называется деполяризацией плазматической мембраны. Смещение
мембранного потенциала в отрицательном направлении называется
гиперполяризацией.
Биофизические основы мембранного потенциала
покоя
Электрические явления в плазматической мембране
определяются распределением ионов между внутренней и внешней сторонами
мембраны. Из химического анализа известно, что концентрация ионов
внутриклеточной жидкости сильно отличается от концентрации ионов во
внеклеточной жидкости. Термин "внеклеточная жидкость" имеет отношение
ко всем жидкостям вне клеток (межклеточное вещество, кровь, лимфа и т.п.). В
таблице представлены концентрации основных ионов в мышечных клетках
млекопитающих и внеклеточной жидкости (миллимоли на литр).
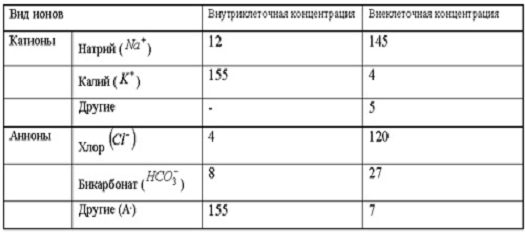
Существуют значительные различия между
концентрацией основных ионов внутри и вне клетки. Внеклеточная жидкость имеет
высокую концентрацию ионов натрия и хлора. Внутриклеточная жидкость имеет
высокую концентрацию калия и различных органических анионов (A-) (заряженные
группы белков).
Различие между концентрациями натрия и калия во
внеклеточной и внутриклеточной жидкостях обусловлены деятельностью
натрий-калиевого насоса, который выкачивает за один цикл 3 иона натрия из
клетки и закачивает 2 иона калия в клетку против электрохимического градиента
указанных ионов. Основная функция натрия-калия насоса - поддержание различия
концентраций ионов натрия и калия по обе стороны плазматической мембраны.
В состоянии покоя проницаемость плазматической
мембраны для ионов калия значительно превышает проницаемость мембраны для ионов
натрия. В нервных клетках соотношения проницаемости соответствующих ионов
составляет 1:0,04.
Этот факт дает возможность объяснять
существование мембранного потенциала покоя.
Ионы калия стремятся покинуть клетку из-за их
высокой внутренней концентрации. При этом перемещения через мембрану
внутриклеточных анионов из-за их больших размеров не происходит. Незначительное
поступление ионов натрия внутрь клетки также не компенсирует выход ионов калия
наружу, так как проницаемость мембраны в покое для ионов натрия мала.
Следовательно, снаружи клетка приобретает дополнительно
положительный заряд и внутри остаётся избыток отрицательного заряда.
Диффузия калия через мембрану - процесс
ограниченный. Ионы калия, проникающие через мембрану, создают электрическое
поле, которое задерживает диффузию других ионов калия. По мере выхода из клетки
калия электрическое поле нарастает и, в конечном итоге, напряжённость достигает
такого значения, когда поток калия через мембрану прекращается. Состояние, при
котором поток ионов по их концентрационному градиенту уравновешивается мембранным
потенциалом, называется состоянием электрохимического равновесия ионов.
Величина такого мембранного потенциала равновесия определяется уравнением
Нернста (при этом считают, что мембрана проницаема только для одного вида
ионов):
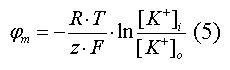
- универсальная газовая постоянная, T -
термодинамическая температура, z - электрический заряд иона, F - постоянная
Фарадея, [K+]i и [K+]o - внутриклеточная и внеклеточная концентрации ионов
калия соответственно.
Вычисления, основанные на уравнении Нернста,
указывают, что внутренняя и внешняя концентрация иона хлора также соответствует
состоянию электрохимического равновесия, но концентрация натрия далека от
равновесия с мембранным потенциалом мембраны.
Уравнение Нернста показывает, что
концентрационный градиент ионов калия определяет величину мембранного
потенциала покоя только в первом приближении. Рассчитанные величины мембранного
потенциала совпадают с экспериментально полученными только при высокой
концентрации калия вне клетки.
Более точная величина мембранного потенциала
покоя может быть вычислена из уравнения Гольдмана-Ходжкина, в котором
учитываются концентрации и проницаемость мембраны для трёх основных ионов
внутри- и внеклеточной жидкостей:
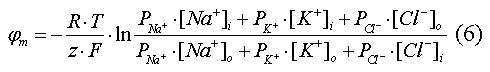
Также в поддержании мембранного потенциала покоя
участвует непосредственно натрий-калий насос, выкачивая три иона натрия из
клетки и закачивая лишь два иона калия. В результате мембранный потенциал покоя
становится более отрицательным, чем был бы, если бы создавался только пассивным
перемещением ионов через мембрану.
Потенциал действия
Если через мембрану нервной или мышечной клетки
проходит кратковременный электрический ток, то мембранный потенциал
подвергается последовательным изменениям, которые специфичны и уникальны для
возбудимых клеток. Возбудимые ткани можно стимулировать также механическими или
химическими средствами, но в экспериментальной работе, как правило,
используются электрические стимулы.
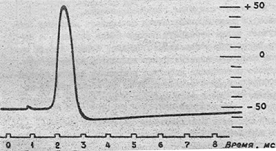
Потенциал действия нервной клетки
На рисунке показан потенциал действия нервной
клетки, записанный с помощью микроэлектрода. Если к клетке прикладывают
кратковременный электрический стимул, мембранный потенциал уменьшается быстро
до нуля.
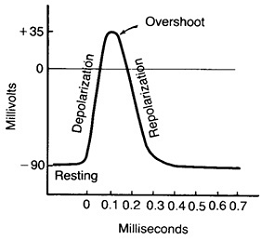
Это отклонение характеризуют как фазу
деполяризации. В течение короткого времени внутренняя среда клетки становится
электроположительна по отношению к наружней (фаза реверсии мембранного
потенциала, или овершут). Затем мембранный потенциал возвращается к уровню
мембранного потенциала покоя (этап реполяризации).
Фазы потенциала действия
Длительность потенциала действия составляет от
0,5 до 1 миллисекунды в больших нервных клетках и несколько миллисекунд в
клетках скелетных мышц. Общая амплитуда - почти 100 - 120мВ, отклонение от
нулевой линии - около 30-50мВ.
Потенциал действия играет ведущую роль в
обработке информации в нервной системе. Он имеет постоянную амплитуду, которая
не является вероятностной величиной. Это имеет большое значение в обработке
информации нервной системой. Кодирование интенсивности раздражения осуществляется
числом потенциалов действия и частотой, с которой потенциалы действия следуют
друг за другом.
электрический заряд поле потенциал
Биофизические основы потенциала действия
Потенциал действия возникает из-за специфических
изменений ионной проницаемости в плазматической мембране. Английский физиолог
Ходжкин показал, что основной механизм потенциала действия состоит в
кратковременном и очень специфическом изменении проницаемости мембраны для
ионов натрия. Ионы натрия при этом поступают в клетку до момента, пока
мембранный потенциал не достигнет потенциала электрохимического равновесия
ионов натрия.
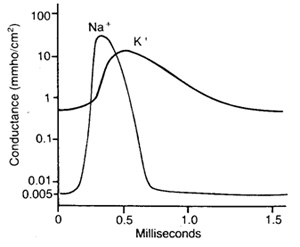
Изменение проницаемости мембраны для ионов
натрия и калия во время потенциала действия.
Проницаемость мембраны для натрия при действии
на клетку электрического стимула возрастает приблизительно в 500 раз и
становится значительно больше, чем проницаемость мембраны для ионов калия. В
клетке резко повышается концентрация ионов натрия. В результате мембранный
потенциал принимает положительное значение, и поток ионов натрия в клетку
замедляется.
Во время возникновения потенциала действия
происходит деполяризация плазматической мембраны. Быстрая деполяризация
мембраны под действием электрического стимула вызывает увеличение её проницаемости
для ионов натрия. Возросшее поступление ионов натрия в клетку усиливает
деполяризацию мембраны, что, в свою очередь, вызывает дальнейшее увеличение
проницаемости мембраны для натрия и т.д.
Но величина мембранного потенциала при
деполяризации не достигает уровня потенциала электрохимического равновесия
ионов натрия. Причиной этому является снижение проницаемости мембраны для ионов
натрия из-за инактивации натриевого трансмембранного переноса. Этот процесс
резко уменьшает проницаемость мембраны для ионов натрия и останавливает наплыв
натрия в клетку.
В этот момент происходит увеличение
проницаемости мембраны для ионов калия, что приводит к быстрому снижению
величины мембранного потенциала к уровню потенциала покоя. Проницаемость
мембраны для ионов калия также снижается до своего нормального значения. Таким
образом, инактивация входящего натриевого тока и повышение проницаемости
мембраны для ионов калия (выходящий ток) ограничивают длительность потенциала
действия и приводят к реполяризации мембраны.
Таким образом, в течение потенциала действия
некоторое количество ионов натрия поступают в клетку. Но это количество
достаточно небольшое. Изменение концентрации ионов в больших нервных клетках
составляет лишь около 1/300000 начальной величины.
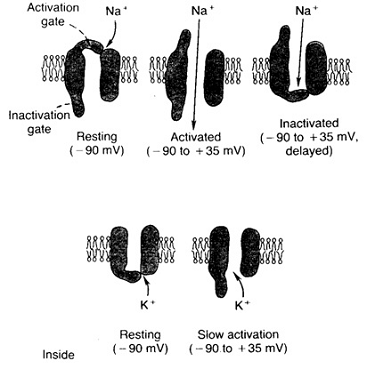
Основной механизм изменений проницаемости
мембраны обусловлен событиями в натриевых и калиевых каналах мембраны.
Состояние их ворот управляется величиной мембранного потенциала. Натриевые
каналы имеют два типа ворот. Один из них, называемые активационными воротами
закрыты в состоянии покоя и открываются при деполяризации мембраны. Поступление
ионов натрия в клетку вызывает открытие всё большего числа активационных ворот.
Второй тип ворот натриевых каналов - инактивационные при усиливающейся
деполяризации мембраны постепенно закрываются, что останавливает приток натрия
в клетку. Деполяризация мембраны также служит причиной открытия дополнительного
числа калиевых каналов, в результате чего увеличивается проницаемость мембраны
для ионов калия и происходит реполяризация мембраны.
Изменение состояния натриевых и калиевых каналов
мембраны в зависимости от величины мембранного потенциала
Распространение потенциала действия
Потенциал действия распространяется вдоль
мембраны нервной и мышечной клеток без уменьшения амплитуды с расстоянием. Этот
процесс обусловлен кабельными свойствами плазматической мембраны, т.е.
способностью проводить электрический ток на небольшие расстояния. Локальный
электрический ток течет в клетку в активной области (где возникает потенциал
действия) и из клетки - в смежной неактивной зоне. Эти ионные токи вызывают
некоторые изменения мембранного потенциала в зоне, прилегающей к месту
возникновения потенциала действия.
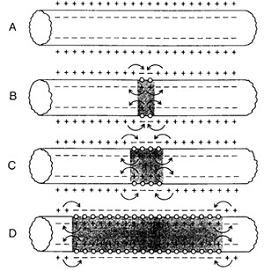
Циклический локальный ток снижает заряд мембраны
в неактивной зоне и деполяризует её. Если деполяризация достигает порогового
уровня, то возрастает проницаемость мембраны для ионов натрия и возникает
потенциал действия. Таким образом потенциал действия распространяется вдоль
нервных и мышечных волокон с постоянной скоростью.
Распространение потенциала действия вдоль
мембраны нервного волокна
Скорость распространения потенциала действия в
нервных волокнах зависит от их диаметра. Она максимальна в наиболее толстых
волокнах, достигая около 100 метров в секунду.
Биопотенциалы
электрические потенциалы в тканях и клетках
(главным образом в клеточных мембранах) живых организмов. Связаны с процессами
возбуждения и торможения у животных и человека и раздражимости у растений. Исследования
биоэлектрических потенциалов применяют с диагностическими целями
(электрокардиография, электроэнцефалография и др.).
Приоритет в открытии «животного электричества»
принадлежит итальянскому врачу и естествоиспытателю Л. Гальвани, описавшему в 1791
сокращение мышцы в ответ на приложение к ней или иннервирующему ее нерву
особого пинцета, одна половина которого состояла из меди, а другая - из железа.
Гальвани объяснял это явление способностью нерва и мышцы быть источниками
электричества, пинцет же, по мнению Гальвани, играл роль проводника,
замыкающего электрическую цепь. В подтверждение своей теории Гальвани приводил
способность некоторых рыб (электрического ската, электрического угря)
вырабатывать электричество и генерировать разряд, подобный молнии, которая была
известна еще в далекой древности.
Мембранная теория биопотенциалов
Первые систематические исследования природы
биопотенциалов и токов в 19 веке принадлежат немецкому электрофизиологу Э.
Дюбуа-Реймону. Измеряя с помощью серебряных электродов разность потенциалов
между поверхностью мышцы (где устанавливался один электрод) и ее внутренней
средой (куда втыкался заостренный второй электрод), он впервые доказал, что в
основе биопотенциалов лежит пространственное разделение положительных и отрицательных
электрических зарядов между наружной и внутренней поверхностью мембраны любой
клетки. В состоянии покоя наружная поверхность клетки всегда заряжена
положительно, а внутренняя - отрицательно, и такой трансмембранный «потенциал
покоя» составляет порядка 0,05-0,09В. Физико-химическую природу потенциала
покоя впервые удалось научно объяснить ученику Дюбуа-Реймона Ю. Бернштейну,
разработавшему в 1903-1911«мембранную теорию биопотенциалов». Опираясь на
данные физикохимии и коллоидной химии о движении ионов в растворах электролитов
и через полупроницаемые органические пленки, а также данные об
электролитическом составе цитоплазмы клетки и внеклеточных жидкостей, Бернштейн
предположил, что мембрана клетки в состоянии покоя не пропускает органические
анионы (которых много внутри клетки) и избирательно проницаема только для ионов
калия, концентрация которых в клетке в 50-100 раз выше, чем в межклеточном
пространстве. Ионы калия диффундируют через мембрану по концентрационному
градиенту наружу, где они скапливаются и придают наружной стороне мембраны
положительный заряд. Одновременно неспособные проходить вслед за калием наружу
органические анионы - противоионы - скапливаются на внутренней поверхности
мембраны и заряжают ее отрицательно. Такой потенциал покоя, возникающий по
разные стороны мембраны, называют диффузионным. В 1930-1940-е мембранная теория
Бернштейна подверглась ревизии. Было показано, что потенциал покоя обусловлен
не только калиевым, но и отчасти натриевым и хлорным диффузионными
потенциалами, и может быть описан с помощью уравнения Гольдмана-Ходжкина-Катца
(см. Потенциал покоя). В 1950-е был установлен новый важный факт, доказывающий,
что сохранение стабильного уровня потенциала покоя клетки требует постоянного
поддержания трансмембранных ионных градиентов калия, натрия и хлора. Как
выяснилось, это происходит за счет работы специальных трансмембранных молекул -
так называемых ионных насосов, трансмембранных сопряженных переносчиков ионов
калия и натрия. Используя энергию АТФ, они постоянно выкачивают ионы натрия из
клетки и закачивают ионы калия внутрь клетки, тем самым поддерживая постоянство
трансмембранных ионных градиентов.
Открытие потенциала действия
Важнейшим этапом в исследовании природы
биопотенциалов в 20 в. стали работы электрофизиологов А. Л. Ходжкина, А. Ф.
Хаксли (Великобритания) и Дж. К. Эклса (Австралия) на гигантских аксонах
кальмара. Используя новый метод фиксации мембранного потенциала с помощью
электронной схемы с отрицательной обратной связью, они впервые прямо
зарегистрировали трансмембранные ионные токи при возбуждении клетки. Было
показано, что возбуждение сопровождается реверсией потенциала (перезарядкой
мембраны): внутренняя поверхность мембраны становится заряженной положительно,
а наружная - отрицательно. Причина перезарядки заключается в том, что в момент
возбуждения мембрана клетки становится кратковременно проницаемой к ионам
натрия, которые, быстро входя в клетку, перезаряжают мембрану. Ходжкин и Хаксли
теоретически, в модельных экспериментах предсказали строение специальных ионных
каналов, способных открываться и пропускать ионы натрия внутрь клетки лишь
кратковременно и в строгой зависимости от величины потенциала на мембране.
Предсказанные свойства молекул натриевых каналов были впоследствии подтверждены
в прямых экспериментах. За раскрытие природы трансмембранных электрических
процессов, происходящих в нервных и других клетках при возбуждении, Ходжкину,
Хаксли и Эклсу в 1963 была присуждена Нобелевская премия. В 1960-1970 годы с
помощью техники введения в клетки тонких стеклянных микроэлектродов (с
диаметром кончика около 0,5 мкм) удалось обнаружить важную роль не только
натриевых, но и кальциевых трансмембранных потоков при возбуждении и генерации
потенциалов действия в нервных и мышечных клетках. В 1980-е годы в работах Э.
Неера и Б. Сакмана был изобретен и впервые использован метод локального
присасывания к мембране клетки с помощью стеклянной пипетки (с диаметром
кончика в несколько микрон), позволивший регистрировать и анализировать ионные
токи, текущие через отдельные ионные каналы.
Таким образом, только в 20 в. стало очевидным,
что «животное электричество», то есть всевозможные биопотенциалы и биотоки
обусловлены движением не электронов, а ионов (натрия, калия, кальция, хлора)
через специальные сложно организованные ионные каналы в мембране, а также
обусловлены диффузией ионов в межклеточной и внутриклеточной средах,
представляющих собой растворы электролитов.
Наряду с потенциалом покоя и потенциалом
действия в 1950-1960 годы были описаны синаптические потенциалы, возникающие в
синапсах и предназначенные для передачи возбуждения от клетки к клетке.
Биоэлектрические потенциалы у растений
Биоэлектрические потенциалы характерны не только
для животных, но и для растений. В 1950-х годах при помощи микроэлектродов, вводимых
в клетку, у нитчатой водоросли нителлы были обнаружены такие же значения
потенциалов покоя, как и у животных клеток - порядка 0,09-0,05 В. Было
установлено, что электрические, механические, химические и другие раздражители
умеренной интенсивности вызывают в местах своего приложения к органам растения
(листу, корню и т. д.) изменения потенциалов, сходные с местными
(подпороговыми) потенциалами у животных клеток. Обнаружены у растений и
специальные потенциалы возбуждения, подобные потенциалам действия животных
клеток. Наиболее приближаются к классическим потенциалам действия электрические
потенциалы, возникающие при распространении волны возбуждения по органам
растения. Так, типичные двухфазные токи действия длительностью 0,1-0,2 мс
сопровождают быстрые движения насекомоядного растения дианова мухоловка, а
также защитную двигательную реакцию складывания листьев у стыдливой мимозы
(Mimosa pudica) в ответ на механическое или электрическое раздражение растения.
Таким образом, биопотенциалы лежат в основе нормальной
жизнедеятельности любой клетки и особенно важны для процессов возбуждения и
торможения у животных и человека и раздражимости у растений. Нарушения
проводимости клеточных мембран могут приводить к серьезным патологиям организма
(вплоть до смерти). Исследования биоэлектрических потенциалов применяют с
диагностическими целями в электрокардиографии, электроэнцефалографии,
электромиографии.
Круговые токи
Законы проведения возбуждения по нервному
волокну
Механизм проведения возбуждения по нервным
волокнам зависит от их типа. Существуют два типа нервных волокон: миелиновые и
безмиелиновые.
Процессы метаболизма в безмиелиновых волокнах не
обеспечивают быструю компенсацию расхода энергии. Распространение возбуждения
будет идти с постепенным затуханием - с декрементом. Декре-ментное поведение
возбуждения характерно для низкоорганизованной нервной системы. Возбуждение
распространяется за счет малых круговых токов, которые возникают внутрь волокна
или в окружающую его жидкость. Между возбужденными и невозбужденными участками
возникает разность потенциалов, которая способствует возникновению круговых
токов. Ток будет распространяться от «+» заряда к«-». В месте выхода кругового
тока повышается проницаемость плазматической мемб-раны для ионов Na, в
результате чего происходит деполяризация мембраны. Между вновь возбужденным
участком и соседним невозбужденным вновь возникает разность потенциалов, что
приводит к возникновению круговых токов. Возбуждение постепенно охватывает
соседние участки осевого цилиндра и так распространяется до конца аксона.
В миелиновых волокнах благодаря совершенству
метаболизма возбуждение проходит, не затухая, без декремента. За счет большого
радиуса нервного волокна, обусловленного миелиновой оболочкой, электрический
ток может входить и выходить из волокна только в области перехвата. При
нанесения раздражения возникает деполяризация в области перехвата А, соседний
перехват В в это время поляризован. Между перехватами возникает разность
потенциалов, и появляются круговые токи. За счет круговых токов возбуждаются
другие перехваты, при этом возбуждение распространяется сальтаторно,
скачкообразно от одного перехвата к другому.
Существует три закона проведения раздражения по
нервному волокну:
) Закон анатомо-физиологической целостности.
) Проведение импульсов по нервному волокну
возможно лишь в том случае, если не нарушена его целостность.
) Закон изолированного проведения возбуждения.
Существует ряд особенностей распространения
возбуждения в периферических, мякотных и безмякотных нервных волокнах:
) В периферических нервных волокнах возбуждение
передается только вдоль нервного волокна, но не передается на соседние, которые
находятся в одном и том же нервном стволе.
) В мякотных нервных волокнах роль изолятора
выполняет миелиновая оболочка. За счет миелина увеличивается удельное
сопротивление и происходит уменьшение электрической емкости оболочки.
) В безмякотных нервных волокнах возбуждение
передается изолированно.
Заключение
Все проявления жизнедеятельности организма
зависят от сложных последовательностей химических реакций, в основе которых
лежит, в частности, явление электричества. Иногда соответствующие процессы
можно изучать, не рассматривая эти силы в явном виде. Такой подход вполне
применим при исследовании, например, регуляции экспрессии генов или механизма
иммунного ответа. Он гораздо менее успешен, когда речь идет о памяти, научении
и регуляции регенеративных процессов. Трудности, с которыми сталкиваются
исследователи, когда пытаются объяснить по крайней мере некоторые биологические
явления - включая саму жизнь - исходя исключительно из биохимических концепций,
заставляют их обратиться к биоэлектрическим факторам. На эту проблему впервые
обратил внимание в 1941 венгерский биохимик А.Сент-Дьёрдьи. Он пришел к выводу,
что феномен жизни нельзя должным образом объяснить просто наличием каких-то
химических веществ: необходимо, чтобы эти вещества находились в определенном
электрическом состоянии. Согласно этой точке зрения, живые и мертвые животные
различаются по своему биоэлектрическому, а не биохимическому статусу. Эти идеи
привели к возрождению интереса к биоэлектричеству.
Список литературы
1. Катц
Б. Нерв, мышца и синапс: Пер. с англ. М., 1968.
. А.Б.
Рубин «Биофизика» 2002-2006г
. Антонов
В.Ф «Биофизика» 2000г