Сезонная динамика параметров замедленной флуоресценции хлорофилла хвойных, произрастающих в районах с разным уровнем загрязнения воздуха
Дипломная
работа
Сезонная динамика параметров замедленной флуоресценции хлорофилла
хвойных, произрастающих в районах с разным уровнем загрязнения воздуха
Введение
Загрязнение окружающей среды
является острой экологической проблемой особенно в городских и промышленных
районах. Воздействие токсикантов приводит к значительному ухудшению состояния и
даже гибели лесов на обширных территориях. Поэтому актуальной задачей является
поиск объективных и оперативных методов ранней диагностики техногенного загрязнения
природной среды.
Проблема загрязнения окружающей
среды чрезвычайно актуальна и для города Красноярска в связи с наличием в
краевом центре крупных предприятий металлургической и химической
промышленности.
Деятельность человека ведет к
необратимым изменениям природной среды, к изменению естественных процессов в
биосфере, регулирующих перераспределение и консервацию различных веществ, в том
числе и вредных.
Непосредственному действию
атмосферных загрязнителей подвергаются наземные органы растения. Из них более
всего поражаются токсическими газами и аэрозолями листья, хвоя и другие зеленые
побеги, которые осуществляют интенсивный газообмен [12].
Вокруг крупных промышленных районов
среди функциональных нарушений растений часто упоминают резкое подавление
деятельности ферментных систем, интенсивности фотосинтеза, повышения
транспирации, усыхание и опадение листьев и хвои, повреждение почек и т.д.
Подобные изменения могут привести к гибели отдельных растений или исчезновению
вида из загрязненного района [58]. Различные исследователи наблюдали видимые
изменения в фотосинтетических структурах: набухание и разрушение хлоропластов,
утолщение и изменение формы тилакоидов, снижение содержания хлорофиллов,
изменение отношения хлорофиллов a/b, другие биохимические изменения [26].
Токсические поллютанты оказывают
разрушающее воздействие на фотосинтетический аппарат и деятельность устьиц, что
является главной причиной ингибирования CO2 - газообмена [15].
В настоящее время в исследованиях
фотосинтеза широко используются методы, основанные на измерении и анализе
замедленной флуоресценции (ЗФ) хлорофилла. Большие перспективы развития данной
области определяются интегральностью флуоресцентных показателей. Это позволяет
использовать параметры флуоресценции для получения разнообразных данных о
функционировании фотосинтетического аппарата растений и изучения действия на
фотосинтез различных факторов [4].
В связи с этим целью данной работы
было изучение сезонной динамики параметров замедленной флуоресценции хлорофилла
хвойных, произрастающих в районах с разным уровнем загрязнения воздуха.
Для достижения этой цели необходимо
решить следующие задачи:
. Изучить влияние загрязнения
воздуха и расположения в кроне на параметры ЗФ хвои сосны обыкновенной и ели
сибирской;
. Рассмотреть возможности
использования сосны и ели в качестве биоиндикаторов в течение года;
. Изучить влияние искусственного
охлаждения побегов сосны в летнее время на параметрах ЗФ хвои.
Работа проводилась на кафедре
Экотоксикологии и микробиологии Сибирского федерального университета под
руководством к.б.н., доцента Н.В. Пахарьковой.
1. Обзор литературы
1.1 Биоиндикационные
методы оценки окружающей среды
биоиндикатор загрязнение
сосна воздух
Метод оценки абиотических и
биотических факторов местообитания при помощи биологических систем называют
биоиндикацией. Организмы или сообщества организмов, жизненные функции которых
тесно коррелируют с определенными факторами среды, называют биоиндикаторами
[1].
В настоящее время в использовании
метода биоиндикации при решении экологических проблем накоплен большой
положительный опыт. Метод широко применяется для оценки состояния и мониторинга
как в природных, так и в нарушенных, в том числе техногенных экосистемах [57].
На сегодняшний день используют
экспресс-методы биоиндикации, которые позволяют на измерении одной или
нескольких функций живого объекта давать быструю и точную оценку по поводу
состояния окружающей среды. В отличие от классических методов, экспресс-методы
более перспективны и менее трудоемки. Преимущество биоиндикационных методов над
физико-химическими заключается в том, что биоиндикация обращается к живому
объекту, который реагирует на весь комплекс загрязняющих веществ.
Виды-биоиндикаторы - датчики широкого спектра. Биоиндикационные методы дают
представление о состоянии живых объектов. Благодаря поглотительной деятельности
растений происходит очищение атмосферного воздуха. Однако, возможности этих
систем ограничены. Растения очень чутко реагируют на загрязнение окружающей
среды, что обуславливает использование их в качестве индикаторов загрязненности
атмосферы [51].
Красноярскими предприятиями
выбрасывается более полутора тысяч различных веществ, среди которых не
маленькая доля ядовитых. Физико-химические методы не в состоянии регистрировать
все влияние на живые системы.
1.2 Основные компоненты
загрязнения атмосферного воздуха
Приземный слой воздуха обычно
содержит, кроме свойственных ему компонентов, примесь различных газов, жидких и
твердых частиц. Они поступают в воздух с земной поверхности в результате человеческой
деятельности, стихийных сил и биологического оборота веществ в биосфере [12].
Многие газообразные соединения, появляющиеся в воздухе в виде примесей, пагубно
действуют на растения. Рассмотрим некоторые из них.
Сернистый ангидрид. (SO2)
образуется при переработке и горении органических веществ (каменный и бурый
угли, нефть и нефтепродукты, древесина), обжиги и плавки серосодержащих руд,
при изготовлении и применении в производственном процессе серной кислоты.
Выбрасывают сернистый ангидрид в большом количестве тепловые электростанции,
предприятия цветной и черной металлургии, коксохимические, цементные,
синтетических волокон и аммиака, целлюлозные, сахароварения и др. [12].тепловая
электростанция, сжигающая в сутки 4500 тонн каменного угля, содержащего 3%
серы, выбрасывает в воздух 270 тонн сернистого ангидрида, а при выплавке меди
из 2250 тонн концентрата руды, содержащего 30% серы, выделяет в атмосферу 1360
тонн сернистого ангидрида [13]. Сернистый ангидрид способен вступать во
взаимодействие с содержащимися в воздухе водяным паром, капельками тумана или
дождя и образовывать сернистую кислоту. Скорость растворения SO2 зависит от концентрации ангидрида в
атмосферном воздухе [33].
Серный ангидрид обладает высокой гигроскопичностью и, соединяясь с водяным паром,
образует аэрозоль серной кислоты, удерживающийся в воздухе в виде тумана.
Серная кислота обладает сильными окислительными свойствами и активно
присоединяет воду. Наличие а атмосферном воздухе совместно сернистого и серного
ангидридов обусловливает их высокую токсичность для растений [47].
Сероводород (H2S) поступает в атмосферу вместе с
другими загрязнителями в относительно не большом количестве по сравнению с
сернистым ангидридом. Он постоянно находится в выбросах коксохимических
предприятий, выделяется при изготовлении искусственного волокна, сахара, в
каменноугольных шахтах, на нефтяных промыслах, нефтеперерабатывающих заводах и
др. сероводород обладает сильными восстановительными свойствами и является
токсичным для растений [47].
Окислы азота и аммиак выбрасывают предприятия, производящие туковые удобрения,
азотистую кислоту и нитраты, анилиновые красители, нитросоединения, вискозный
шелк, целлулоид, фотопленку, а так же выхлопные газы автотранспорта. Окислы
азота (NO2, N2O3, N2O5) легко растворяются в воде, содержащейся а воздухе, образуя
аэрозоль азотной и малоустойчивой азотистой кислот [47].
Аммиак (NH3)
выделяется в атмосферу в небольших количествах при производстве аммонийных
удобрений, мочевины, азотной кислоты, на сахарных, кожевенных и др.
предприятиях. Аммиак обладает восстановительными свойствами со слабовыраженной
токсичностью [47].
Фтористые соединения являются одними из наиболее вредных для растений. Фтор легко
вступает в реакцию почти со всеми элементами. Поэтому в свободную атмосферу он
поступает не в чистом виде, а в соединении с другими веществами в виде газа или
пылевидных частиц. Источниками загрязнения воздуха фтористыми веществами
являются алюминиевые и криолитовые заводы, предприятия, производящие фосфаты и
фосфорные удобрения, эмалевые и керамические изделия и т.д. некоторое
количество фтористых соединений выделяется из топок при сжигании угля и из
флюсов, применяемых при выплавке чугуна. Газообразные или растворенные в воде
соединения фтора проникают в растения через листья и корни. Фтористые
соединения являются высокотоксичными для растений. Не редки случаи полной
гибели листьев деревьев у заводов. Губительное действие фтора на растения
усиливается при высокой влажности воздуха и моросящих осадках [47].
Соединения хлора являются
высокореакционными окислителями. Загрязнение атмосферного воздуха хлористыми
соединениями происходит от титано-магниевых заводов, химических предприятий,
производящих инсектициды и гербициды, соляную кислоту, органические красители,
цемент, суперфосфат, уксусную кислоту гидролизный спирт, хлорную известь, соду
и др. хлористый водород и другие его соединения необходимы растениям в
ограниченном количестве. В случае попадания в организм в высоких дозах хлор
вызывает глубокие структурные и функциональные нарушения, нередко ведущие к
гибели растений [47]
Окись углерода выделяется всюду при неполном сгорании веществ, содержащих
углерод (каменный уголь, нефтепродукты, природный газ). Окись углерода является
восстановителем. Она отрицательно влияет на растения при сравнительно высокой
концентрации - свыше 1% [47].
Аэрозоли бывают твердые и жидкие. Под аэрозолями понимают содержащиеся в
воздухе, как дисперсионной среде, мельчайшие твердые и капельно-жидкие частицы
(диспергенты) во взвешенном состоянии. Совокупность твердых частиц проявляется
в виде дыма, а жидких - тумана или облака. Размер частиц аэрозолей колеблется в
широких пределах - от сотых долей микрона до десятков и сотен микрон. Частицы
менее 5 мк могут длительное время находиться в воздухе в виде взвесей, а при
достижении размера более 10 мк они быстро оседают. Основными источниками
выброса твердых и жидких частиц в атмосферу являются тепловые электростанции,
сжигающие бурый уголь, металлургические предприятия, цементные и сажевые
заводы, а так же транспорт и другие. При изготовлении каждой тонны цемента
завод теряет 240 кг продукции в виде пыли. Жидкие частицы аэрозоля образуются
при конденсации паров кислот, оснований, фенолов, смол и иных веществ, а так же
при химическом реагировании загрязнителей с водяным паром или капельками воды,
содержащимися в атмосфере при тумане или дожде. Так, в насыщенном влагой
воздухе образуются сернистая серная, азотная, соляная, фторидная и другие
кислоты из соответствующих ангидридов, поступивших в атмосферу в виде газа [47].
Древесные растения, обладающие
огромной листовой поверхностью, служат хорошим фильтром для очищения воздуха от
аэрозолей. Твердые и жидкие частицы, соударяясь с листьями, оседают на них и
смываются жидкими осадками [47].
Указанные вещества в момент поступления
в атмосферу или в ночное время не оказывают повреждающего или раздражающего
действия на живые организмы. Их токсичность резко возрастает после некоторого
периода освещения солнечными лучами [47].
Ассимиляционные органы растений
первыми и в наивысшей степени повреждаются токсическими веществами,
содержащимися в воздухе промышленных районов и крупных городов. Это вызвано
тем, что листья по своему строению функциональной роли приспособлены к более
интенсивному газообмену по с равнению с другими органами растения. Появление
внешне различимых признаков повреждения растения газами или аэрозолями
свидетельствует о происшедших в организме глубоких, как правило, необратимых
изменениях, заканчивающихся разрушением пигментов, клеточных и субклеточных
структур [47].
Растения делят по степени
противодействия организма повреждающему агенту на устойчивые, среднеустойчивые
и нестойкие или по степени поражения - на нечувствительные,
среднечувствительные и очень чувствительные [47].
1.3 Сосна обыкновенная
как биоиндикатор загрязнения окружающей среды
Вредное влияние загрязненного
воздуха на растения происходит как путем прямого действия газов на
ассимиляционный аппарат, так и в результате косвенного влияния через почву. При
чем прямое действие кислых газов приводит к отмиранию отдельных органов
растений, ухудшение их роста и продуктивности. Трудно дать полную
характеристику всех вредных поллютантов, так как их количество и удельный вес в
загрязненном воздухе быстро изменяются. В результате взаимодействия веществ образуются
новые соединения, нередко даже более токсичные, чем исходные компоненты [27].
При воздействии различных доз
токсикантов на растения можно выделить четыре типа повреждений, отличающихся по
характеру и глубине расстройства метаболизма: острая, капельно-ожоговая,
кумулятивное и скрытое. Острые повреждения вызываются воздействием высоких
концентраций поллютантов. Поражение развивается в течение нескольких часов
(дней) и проявляются в виде хлороза с последующей некротизацией тканей. Кислый
газ, поступая в клетку через устьица или кутикулу, в значительных количествах
накапливается в хлоропластах и приводит к подавлению фотосинтеза [19].
Капельно-ожоговые в отличие от
острого поражения, охватывающего всю хвоинку и возникающего по всей кроне или
большей ее части, проявляется в виде некрозных точек и более крупных пятен,
перепоясывающих хвоинки и постепенно расширяющихся, и чаще всего образуется при
высокой влажности воздуха [8, 33].
Кумулятивное повреждение развивается
при длительном действии низких концентраций токсикантов. Для этого типа
повреждений характерно стойкое нарушение основных метаболических процессов,
внешне выражающееся в уменьшении размеров и массы хвои, преждевременном их
опадании, изреженности крон деревьев, депрессии роста, ранним старением. Кумулятивное
повреждение со временем приводит к избыточному накоплению токсикантов
ассимиляционных органах и проявлению некрозов. Выше перечисленные типы
повреждений определяются визуально или с помощью морфологического анализа [8,
33].
Скрытые повреждения вызываются очень
низкими концентрациями токсикантов, действующими постоянно или переодически. К
таким повреждениям относятся нарушения физиолого-биохимических процессов
(увеличение проницаемости мембран, изменение активности ряда ферментов,
ингибирование фотосинтеза и так далее) накопление скрытых повреждений приводит
к снижению роста и продуктивности растений, потере резистентности к
климатическим факторам, патогенным микроорганизмам и насекомым. Именно на
стадии скрытых повреждений необходимо наладить контроль за состоянием растений,
подверженных к действию промышленного загрязнения воздуха, что может быть
достигнуто с помощью методов флуоресцентного анализа [2, 4].
Повышенная чувствительность хвойных
связана с длительными сроками жизни хвои и поглощения газа, слабым развитием
запасающих тканей и, соответственно, недостаточными накоплениями резервных
веществ, не высокой регенеративной способностью. Тем не менее, разные виды
хвойных обладают различными биоиндикационными возможностями. Одна из наиболее
важных характеристик - сезонный ритм ростовых процессов, в частности сроки
начала и конца вегетации. Деревья и кустарники, рано начинающие и рано
заканчивающие вегетационный цикл особенно сильно повреждаются в условиях
загрязнения. Виды, начинающие вегетировать в более поздние сроки и поздно
заканчивающие вегетацию, наиболее устойчивы к загрязнению.
Сосна обыкновенная (Pinus sylvestris) - одна из важнейших
лесообразующих пород, занимающая по численности треть хвойных. Своеобразие
климата Сибири предполагает высокую адаптацию к температурным условиям [30].
Одним из преимуществ данного объекта
исследования является возможность использования его в течении всего года. Сосна
сохраняет фотосинтетическую активность при низких температурах, квантовый выход
не меняется.
Для хвойных характерны сезонные
изменения содержания и соотношения хлорофилла, а так же каротиноидов. От начала
формирования молодой хвои, количество хлорофилла а, хлорофилла b и сумма каротиноидов в ней, плавно
нарастает с достижением максимума в августе - сентябре. Следует отметить, что
больше снижается содержание хлорофилла b, что приводит к увеличению отношения
хлорофиллов a/b [54].
Отношение хлорофиллов a/b в течение
большей части года изменяется в противофазе с содержанием хлорофилла b, и
только в сентябре происходит одновременное снижение как содержание хлорофилла
b, связанного с компонентами фотосистемы 1 (ФС 1) и фотосистемы 2 (ФС 2)
значительно превышает такую же для хлорофилла светособирающего комплекса (ССК).
Известно, что хлорофилл b сосредоточен преимущественно с ССК и ФС 2. скорость
разрушения пигментов не одинакова в разные периоды года. Начинаясь осенью,
процесс деградации хлорофилла развивается к зиме и затормаживается, не смотря
на морозы, в январе - феврале. Процесс деградации у хвои протекает при
установлении холодной и солнечной погоды зимой. В конце весны с началом нового
вегетационного периода у хвои начинается активная регенерация фонда обоих
компонентов хлорофилла, и как следствие, постепенное восстановление окраски
хвои в июле [54].
Метод регистрации ЗФ хлорофилла у
такой древесной породы, как сосна обыкновенная, обеспечивает надежное выявление
начальных стадий повреждения растений при загрязнении атмосферы [6].
1.4 Ель сибирская как
биоиндикатор загрязнения окружающей среды
Известна высокая чувствительность
хвойных растений к различным видам загрязнений, что обуславливает их широкое
использование в качестве биоиндикаторов при оценке качества окружающей среды
[33].
Повышенная чувствительность хвойных
связана с длительными сроками жизни хвои и поглощения газа, слабым развитием
запасающих тканей и, соответственно, недостаточными накоплениями резервных
веществ, не высокой регенеративной способностью. Тем не менее, разные виды
хвойных обладают различными биоиндикационными возможностями. Одна из наиболее
важных характеристик - сезонный ритм ростовых процессов, в частности сроки
начала и конца вегетации. Деревья и кустарники, рано начинающие и рано
заканчивающие вегетационный цикл особенно сильно повреждаются в условиях
загрязнения. Виды, начинающие вегетировать в более поздние сроки и поздно
заканчивающие вегетацию, наиболее устойчивы к загрязнению.
Ель сибирская (Picea obovata) - одна из важнейших
лесообразующих пород, занимающая по численности треть хвойных. Своеобразие
климата Сибири предполагает высокую адаптацию к температурным условиям [30].
Одним из преимуществ данного объекта
исследования является возможность использования его в течении всего года. Ель
сохраняет фотосинтетическую активность при низких температурах, квантовый выход
не меняется.
Для хвойных характерны сезонные
изменения содержания и соотношения хлорофилла, а так же каротиноидов. От начала
формирования молодой хвои, количество хлорофилла а, хлорофилла b и сумма каротиноидов в ней, плавно
нарастает с достижением максимума в августе - сентябре. Следует отметить, что
больше снижается содержание хлорофилла b, что приводит к увеличению отношения
хлорофиллов a/b [54].
Отношение хлорофиллов a/b в течение
большей части года изменяется в противофазе с содержанием хлорофилла b, и только
в сентябре происходит одновременное снижение как содержание хлорофилла b,
связанного с компонентами фотосистемы 1 (ФС 1) и фотосистемы 2 (ФС 2)
значительно превышает такую же для хлорофилла светособирающего комплекса (ССК).
Известно, что хлорофилл b сосредоточен преимущественно с ССК и ФС 2. скорость
разрушения пигментов не одинакова в разные периоды года. Начинаясь осенью,
процесс деградации хлорофилла развивается к зиме и затормаживается, не смотря
на морозы, в январе - феврале. Процесс деградации у хвои протекает при
установлении холодной и солнечной погоды зимой. В конце весны с началом нового
вегетационного периода у хвои начинается активная регенерация фонда обоих
компонентов хлорофилла, и как следствие, постепенное восстановление окраски
хвои в июле [54].
Ель сибирская занимает почти всю
северную половину европейской части нашей страны. Ель теневыносливая и
влаголюбивая порода. Ель очень чувствительна к весенним заморозкам. Пигментный
состав фотосинтетического аппарата ели более чувствителен к действию
загрязнителей, по сравнению с другими хвойными [30].
.5 Состояние растений,
произрастающих в районах с различной степенью техногенного загрязнения
По валовому выбору вредных веществ,
Красноярский край прочно занимает первое место среди республик, краев и
областей. Среди отраслей существенный вклад в загрязнение атмосферы воздуха
вносит: цветная металлургия, химическая промышленность, электроэнергетика,
жилищно-коммунальное хозяйство, целлюлозно-бумажный комбинат. Наиболее больший
объем вредных веществ выбрасывает: АО «Красноярский алюминиевый завод»,
Красноярская ГРЭС-2, ТЭЦ-1, ТЭЦ-2. велика доля автотранспорта в суммарных
выбросах в Красноярске [57].
Вокруг крупных промышленных районов
среди функциональных нарушений растений часто упоминают резкое подавление
деятельности ферментных систем, интенсивности фотосинтеза, повышения
транспирации, усыхание и опадание листьев, повреждение почек и т.д. подобные
изменения могут привести к гибели отдельных растений или исчезновение вида из
загрязненного района. Испытания показали, что лиственные породы более устойчивы
к загрязнению воздуха, чем хвойные, пылеулавливающая способность их выше [58].
Токсичные поллютанты оказывают
разрушающее воздействие на фотосинтетический аппарат и деятельность устьиц, что
является главной причиной ингибирования процесса СО2 - газообмена. В то же время деятельность воздушных
загрязнителей приводит к деструкции корневой системы и существенным изменениям
в структуре проводящей ксилемы, вследствие чего возникают нарушения в водном
обмене деревьев. Усиливающийся водный стресс является дополнительной причиной
значительного снижения фотосинтеза [15].
В районах, в которых в течение
нескольких лет происходит выброс SO2 и HF, у деревьев поражает
листья и изменяет режим роста. Эти фенотипические изменения приводят, в
основном, к снижению нормальной скорости роста, вызывают деформацию ствола,
веток и кроны деревьев. Первыми изменениями реакции растения на загрязненную
среду является снижение морозоустойчивости деревьев, характеризуется краевым поражением.
Наблюдается усиление повреждения хвойных зимой [35].
Изменение фотосинтетического
аппарата под действием атмосферных загрязнений определяется высокой реакционной
способностью ХБК и переносчиков электрон-транспортной цепи (ЭТЦ) и
стабильностью мембранных структур хлоропластов. Наблюдаемое при этом подавление
фотосинтеза сопровождается трансформацией биохимических процессов и пигментного
аппарата [56].
Загрязнение воздуха снижает
содержание хлорофилла как в ССК, так и в комплексах ФС1 и ФС2, приводит к
возрастанию внешнего максимума отношения хлорофилла a/b, что свидетельствует о большой
чувствительности пораженной хвои к влиянию зимнего стресса. Загрязнение воздуха
помимо непосредственного повреждения фотосинтетического аппарата может влиять
на эффективность адаптации его к низким температурам в период покоя и снижает
его устойчивость к повреждениям в зимний период [5].
Хвойные относительно устойчивы в
раннем периоде роста и становятся очень чувствительными при пике роста
древесины, при чем эта чувствительность сохраняется до ранней стадии зрелости
[35].
Атмосферное загрязнение как фактор
стресса в окружающей среде связано со многими другими факторами стресса.
Загрязняющие вещества снижают устойчивость растений к морозу, зимним условиям,
засухе. Фториды обычно вызывают краевые некрозы листьев или некрозы кончиков
игл хвойных [35].
При рассмотрении растений с точки
зрения концентрации загрязненных веществ в атмосфере, было отмечено, что
растения более устойчивы зимой, чем в вегетационный период. Первыми изменениями
реакции растений на окружающую среду является снижение морозоустойчивости
растений, характеризуется краевым поражением. Наблюдается усиление повреждения
хвойных зимой. Под действием загрязненного воздуха меняется состав сахаров в
иглах [9].
Аккумуляция загрязняющих веществ на
зеленую поверхность зимой вызывает ее разрушение. Быстрее всего этот процесс
протекает в январе и феврале. Зимой хвойные обычно толерантны к более низким
температурам, чем те, которые имеют место в действительности, а зимние
повреждения могут быть связаны с зимней засухой или токсичными свойствами
аккумулированных загрязняющих веществ, влияющих на мембраны фотосинтетического
аппарата [61].
1.6 Состояние покоя у
древесных растений
Одно из приспособлений древних
растений к переживанию неблагоприятных условий зимнего периода - их способность
переходить в состояние зимнего покоя.
Физиологическое значение покоя
заключается в приспособлении к неблагоприятным условиям, защите от
обезвоживания (апексы побегов прекращают активный рост, покрываются чешуями,
образуя зимние покоящиеся почки) [35].
У растительных организмов периоды
вегетации, интенсивного роста чередуются с покоем. В состоянии глубокого покоя
у растений резко заторможен обмен веществ и прекращается видимый рост.
Состояние зимнего покоя период
интенсивной деятельности меристемы - образовательной ткани, как у вечнозеленых,
так и у листопадных растений в это время закладываются зачатки листьев в
вегетативных почках и элементов цветков - в цветочные. Без этого весной переход
к активной жизнедеятельности был бы невозможен. Таким образом, покой -
обязательное условие дальнейшего нормального роста а период вегетации [51].
Способность погружаться в состояние
покоя, выработанная растениями в ходе эволюции - важное приспособление к
периодическому наступлению неблагоприятных внешних условий. В состоянии
вынужденного покоя растения находятся в конце зимы. Октябрь-ноябрь - глубокий
покой, который не могут нарушить самые благоприятные для роста условия.
Глубокий покой - необходимая фаза развития растений, сменяющая период вегетации
[35].
Продолжительность периода покоя у
разных деревьев и кустарников различна. Самый длительный покой у ели сибирской,
сосны обыкновенной. В зимний период происходит ряд изменений в организации и
функционировании фотосинтетического аппарата хвои. Наблюдается пожелтение хвои,
обусловленное фотоокисление хлорофилла, при этом сначала происходит деградация
фонда пигментов ССК, за счет более высокомолекулярных олигомеров
соответствующих ХБК. У пожелтевшей хвои изменяется структура хлоропластов,
снижается активность фотосинтетического аппарата, что проявляется в снижении
активности ФС1 и ФС2, в замедлении переноса электронов на уровне пластохинона.
Изменение активности фотосинтетического аппарата в зимний период обратимы, т.к.
деревья хвойных пород приспособлены к переживанию морозов и сохраняют иглы в
течение нескольких лет [20].
Подготовка фотосинтетического
аппарата происходит осенью при переходе деревьев в состояние покоя. В этот
период при положительных и небольших отрицательных температурах подавляется
переменная флуоресценция хлорофилла в хвое сосны и изменяется температурная
зависимость выхода флуоресценции на уровне F0 в хвое, а также в феллодерме коры у различных пород: max около +50°С
подавляется, а появляется новый max около +80°С [3].
При нагревании хлорофиллсодержащих
тканей выход постоянной флуоресценции возрастает при температуре около +45°С в
следствие тепловой инактивации РЦ ФС2, после чего уменьшается из-за денатурации
ССК. В результате на термограмме постоянной флуоресценции возникает max около +50°С, при более
высокой температуре иногда обнаруживается второй max около +65°С, который
связывают с тепловой инактивацией ХБК ФС2. переход растений в состояние покоя
сопровождается изменением термограммы флуоресценции, max около +55°С исчезает и
резко возрастает интенсивность флуоресценции при температуре выше +65°С. Подавление
низкотемпературного max с усилением безизлучательного тушения возбуждения в ФС2, которое
проявляется, например, при фотоингибировании фотосинтеза. При переходе растения
в состояние покоя чувствительность их к фотоингибированию может вырасти, т.к.
при подавлении синтеза белков замедляется восстановление активности
фотоингибирующих РЦ ФС2. появление интенсивной флуоресценции при высокой
температуре нельзя связывать с усилением флуоресценции ФС1, т.к. возрастание
количества ФС1 в состоянии покоя представляется маловероятным. Появление
высокотемпературной флуоресценции в покоящихся тканях, может быть обусловлено
накоплениями в клетках веществ, способствующих такому переходу, либо изменении
состояния самих тилакоидных мембран [53].
Причиной высокотемпературного max может быть «разгорание»
флуоресценции более термостабильного ХБК ФС1 при инактивации ее РЦ.
Определенный вклад в появлении
высокотемпературного пика может дать ХБК ФС2 при условии ее локализации в
межгранных участках мембран тилакоидов и дефиците СХБК [5].
В настоящее время накоплено много
данных о том, что состояние покоя контролируется с помощью гормонов. Вхождение
в покой связано с увеличением содержания абсцизовой кислоты (АБК), а выход из
покоя с появлением цитокининов и гиббереллинов [50].
АБК - тормозит рост проростков,
подавляет открытие устьиц, активизирует отток метаболитов из листа и ингибирует
процессы фотосинтеза, способствует развитию состояния покоя, тормозит синтез
гидролитических ферментов (амилазы, протеиназы), необходимых для различных
процессов вегетации растений [32].
Важную роль в переходе растения в
состояние покоя играет и этилен, вызывающий старение листьев, определяющий
пребывание почек в состоянии покоя. Одновременно в тканях растений уменьшается
содержание естественных стимуляторов роста.
На переход растений в состояние
покоя оказывает влияние и температура. Для некоторых видов понижение ночных
температур - главный сигнал к покою. Также сигналом к покою служит уменьшение
продолжительности светового дня [51].
Из литературных данных известно, что
максимальная скорость фотосинтеза в интервале +20 - 25°С. Нижний предел
температур, при котором еще наблюдается фотосинтез, колеблется от +15°С (сосна,
ель) до +3°С, у большинства высших растений фотосинтез прекращается
приблизительно при 0°С. Устойчивость растений к низким температурам создается
благодаря внутриклеточным изменениям. Роль антифриза - веществ, которые снижают
температуру замерзания растворов, находящихся в клеточных вакуолях, играют
сахара: они накапливаются в клеточном соке во время подготовки растения к зиме.
Они же предохраняют белки цитоплазмы от коагуляции при понижении температуры.
Чем больше сахаров накапливает растение а своих тканях, тем лучше они
противостоят действию низких температур [51].
Под действием загрязнения воздуха
меняется состав сахаров в листьях и иглах, например, обнаружено осеннее
снижение содержания глюкозы и сахара в загрязненной хвое ели европейской по
сравнению с незагрязненной [59].
Нарушается транспирация: стадия
полного покоя развивалась не полностью, загрязненные иглы были готовы к
транспирации в январе [59].
При выходе растения из состояния
покоя наблюдается появление при +45 - 55°С низкотемпературного пика, что
связано с тепловым повреждением мембран тилакоидов на уровне ХБК ФС2 и СХБК, затрагивающим
целостность РЦ и структурно функциональную связь между ФС2 и хлорофилл a/b белковым комплексом. Возрастание
выхода флуоресценции при +70 - 80°С связано с выходом хлорофилла в липидную
фазу при денатурации ХБК.
.7 Замедленная
флуоресценция и ее использование для оценки состояния растения
Флуоресценция
хлорофилла
Фотосинтез зеленых растений очень
чувствителен к различным изменениям факторов внешней среды. Сначала прошлого
века его все чаще используют с помощью постоянно расширяющегося арсенала методов
для определения реакции растений на стрессы, в особенности на загрязнение
воздуха [1].
Фотосинтез - стабильное
фундаментальное явление, поэтому, снижение его интенсивности свидетельствует об
ухудшении состояния растения. Флуоресценция выступает как инструмент
исследования фотосинтеза.
Характер изменения
первичных стадий фотосинтеза непосредственно отражается в изменении
флуоресценции хлорофилла в фотосинтетических мембранах клеток. Для понимания
этой взаимосвязи достаточно напомнить, что поглощение кванта света переводит
молекулу хлорофилла в электронное возбужденное состояние, энергия которого в
растворе при отсутствии фотосинтеза переходит либо в тепло, либо в
флуоресценцию. В фотосинтетической мембране энергия электронного возбуждения
хлорофилла используется в реакционных центрах (РЦ) для генерации потока
электронов в первичных стадиях фотосинтеза, необходимых для восстановления НАДФ
и образования АТФ. Напомним, что первичные процессы фотосинтеза высших растений
осуществляются при участии двух фотосистем, функционирующих последовательно.
Фотосистема 2 разлагает воду с выделением свободного кислорода и отдает
электрон через цепь переносчиков на фотосистему 1, которая уже восстанавливает
НАДФ. В клетке в основном флуоресцирует хлорофилл, принадлежащий фотосистеме 2,
и именно изменения его флуоресценции говорят о состоянии реакционных центров
этой фотосистемы. При активном фотосинтезе, когда все РЦ находятся в открытом
рабочем состоянии, в условиях слабого освещения почти вся поглощенная энергия
света используется в процессе фотосинтеза. Поэтому интенсивность флуоресценции
хлорофилла в клетке намного ниже, чем в растворе.
Однако и здесь
небольшая часть энергии электронного возбуждения (не более 3%) переходит в
энергию света флуоресценции в виде так называемой фоновой флуоресценции F0. Как правило, в
нормальных условиях величина F0 мала, что говорит об активном использовании клетками энергии
поглощенного света. Но если при каких-либо воздействиях нарушается состояние
фотосинтетических мембран, то центры (РЦ) переходят в неактивное (закрытое)
состояние, когда происходит прекращение потока электронов в первичных процессах
фотосинтеза. В этих условиях поглощенная энергия света уже не может
использоваться в фотосинтезе, поэтому и флуоресценция хлорофилла возрастает.
Можно полностью вывести из рабочего состояния РЦ, например, при действии
ингибитора потока электронов диурона. В этом случае флуоресценция сильно
возрастает и приближается к своим максимальным значениям Fт. Заметим, что закрыть
центры можно создавая также избыточную освещенность клеток, когда происходит
световое насыщение фотосинтеза. Фотосинтетическая цепь переноса электрона как
бы захлебывается от избытка поглощенной световой энергии, переводя все большую
часть поглощенной энергии света в флуоресценцию.
Можно найти разницу между
интенсивностями флуоресценции хлорофилла при закрытых и открытых РЦ (Fv = Fт - F0), которую
называют переменной флуоресценцией (Fv) хлорофилла в
клетках. Как видно, величина Fv соответствует той части энергии света, которая используется
открытыми реакционными центрами в фотосинтезе, то есть может характеризовать
активность начальных стадий фотосинтеза. На практике оценивают отношение Fv./Fт, величина которого тесно связана с первичной продуктивностью
фитопланктона в природных водоемах. Она хорошо коррелирует с фотосинтетической
продукцией клеток, определенной классическими методами по восстановлению СО2
с помощью радиоактивных изотопов 14С [36].
Удобным и перспективным
биофизическим методом, несущим информацию о функционировании первичных реакций
фотосинтеза является метод регистрации замедленной флуоресценции.
Явление замедленной флуоресценции
наблюдается у всех видов фотосинтезирующих растений. Замедленная флуоресценция
возникает при излучательной дезактивации первого синглетного возбужденного
состояния хлорофилла и обусловлена вторичным возбуждением хлорофилла, а при
обратных реакциях образования на свету фотопродуктов, в результате рекомбинации
зарядов первичного акцептора электронов и хлорофилла реакционного центра
фотосистемы 2 [4].
Замедленная флуоресценция зеленых
растений возникает в основном в реакционном центре фотосистемы 2, однако,
чрезвычайно слабая замедленная флуоресценция может наблюдаться в фотосистеме 1.
Интенсивность замедленной
флуоресценции пропорциональна скорости реакции зарядов в реакционном центре
фотосистемы 2, пропорционально концентрации центров с разделенными зарядами.
Концентрация последних зависит от
скорости последующих стадий переносов электронов. Именно этим объясняется
зависимость интенсивности и других параметров замедленной флуоресценции от
изменения скорости электронного транспорта в хлоропластах при изменении внешних
условий.
Использование
замедленной флуоресценции для оценки состояния растений
Изменения параметров замедленной
флуоресценции при действии факторов внешней среды можно использовать для
определения устойчивости растений. Максимальная амплитуда неблагоприятного
фактора, при которой еще нет существенных изменений параметров замедленной
флуоресценции, будет характеризовать устойчивость фотосинтетического аппарата
растений к этому фактору. Во многих случаях устойчивость фотосинтетического
аппарата, регистрируемая методом замедленной флуоресценции, коррелирует с общей
устойчивостью растительного организма к тому или иному фактору [24].
Преимущество флуоресцентных методов
исследования заключается в том, что информацию о содержании хлорофилла,
организации фотосинтетического аппарата и его активности можно получить за
очень короткий отрезок времени, как при контактном, так и бесконтактном способах
измерения, что очень важно для решения экологических проблем.
Методы, основанные на флуоресцентных
параметрах, характеризуются высокой чувствительностью, возможности
автоматизации полученной информации, проведения ранней диагностики [33]. Методы
дают нам то, что ошибка в самом плохом случае не превышает 10%, в идеале она
может быть равной 1%.
Таким образом, благодаря
использованию метода замедленной флуоресценции мы можем с высокой
оперативностью и чувствительностью получать многообразную информацию о
фотосинтетическом аппарате, по которой можно судить о влиянии стрессовых
факторов на исследуемый объект.
2. Материалы и методы
В качестве объектов исследований
использовались деревья сосны обыкновенной (Pinus sylvestris) и ели сибирской (Picea obovata) в возрасте от 20 до 35
лет, входящих в состав искусственных насаждений г. Красноярска, на восьми
пробных площадках.
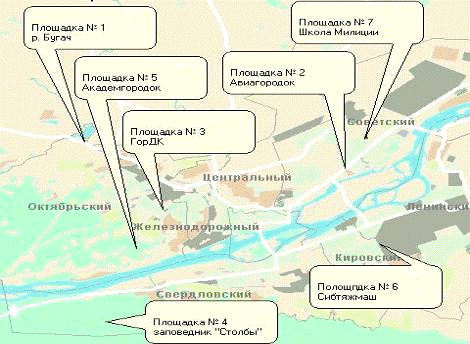
Площадка №1 (Октябрьский район) - расположена в городе, вблизи Красноярского
маргаринового завода. Поток автотранспорта средний. Деревья визуально не
повреждены. Хвоя не запылена. Виды: ель, сосна.
Площадка №2 (Советский район) - расположена в городе, на пересечении улицы
Краснодарская и проспекта Металлургов, испытывает сильный поток автотранспорта.
Деревья визуально не повреждены. Хвоя запылена. Виды: ель.
Площадка №3 (Октябрьский район) - расположена в городе, на пересечении
проспекта Свободный и проспекта Октябрьский (остановка ГорДк). Испытывает
влияние большого потока автотранспорта и выбросов промышленных предприятий,
котельных. Деревья визуально не повреждены. Хвоя запылена. Виды: ель.
Площадка №4 (Свердловский район) - заповедник «Столбы». Расположена за
городом, в трех километрах от дороги. Деревья визуально не повреждены. Хвоя не
запылена. Виды: ель, сосна.
Площадка №5 (Октябрьский район) - расположена в городе, в Академгородке
недалеко от дороги. Испытывает влияние сильного потока автотранспорта. Деревья
визуально не повреждены. Хвоя не запылена. Виды: ель, сосна.
Площадка №6 (Кировский район) - расположена в городе, вблизи завода СибТяжМаш
по улице Мичурина. Деревья визуально не повреждены. Хвоя не запылена. Виды:
ель, сосна.
Площадка №7 (Советский район) - расположена в городе, рядом со Школой
милиции, испытывает влияние сильного потока автотранспорта. Деревья визуально
не повреждены. Хвоя запылена. Виды: ель, сосна.
Площадка №8 (д. Погорелка) - расположена на расстоянии 35 километров от
Красноярска. Деревья находятся в 1,5 км от трассы, визуально не повреждены,
хвоя не запылена. Площадка является контрольной. Виды: ель, сосна.
Для исследования на каждой площадке
было выбрано по три дерева, примерно одного возраста, визуально не поврежденных
и в одинаковых условиях освещенности. От каждого дерева брали по 4 побега. Для
исследований брали хвою первого и второго года жизни.
Измерения проводили сразу после
сбора, после выведения растений из состояния покоя, а так же после экспозиции в
лабораторных условиях. Для оценки низкотемпературного стресса, хвою с каждой
площадки держали по одной неделе при комнатных условиях (+24º С), при отрицательных (-
16,2º и -2º С) температурах.
Регистрацию ЗФ проводили на
флуориметрах «Фотон-9» и «Фотон-10», разработанных на кафедре экотоксикологии и
микробиологии под руководством проф. Ю.С. Григорьева.
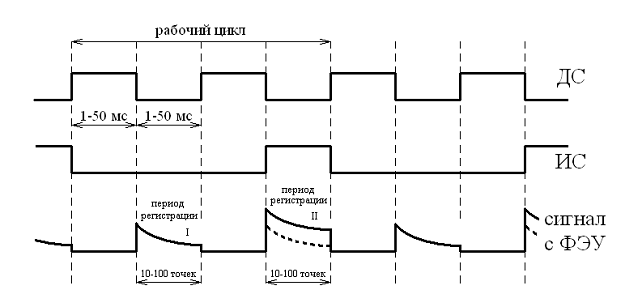
Рис. 1. Диаграмма работы прибора
Фотон 9
Обозначения:
ДС - действующий (яркий, красный)
свет, излучаемый светодиодами
ИС - измерительный (слабый, синий)
свет, излучаемый светодиодами (зонд)
ЗФ - замедленная флуоресценция,
регистрируемая после вспышек ДС
Ф - флуоресценция (быстрая),
возбуждаемая ИС и регистрируемая в период его включения
ФЭУ - фотоэлектронный умножитель
Рабочий цикл включает два импульса
ДС и два темновых промежутка между ними, в один из которых подается импульс ИС.
Функционирование обоих приборов
сходно. В основу работы прибора положен принцип измерения послесвечения
хлорофиллсодержащего растительного объекта, в промежутках между импульсами
возбуждающего света (рис. 1).
По своей природе это свечение
является фотохемилюминесценцией хлорофилла, которое благодаря идентичности
спектру излучения быстрой флуоресценции (фотолюминесценции), получило название
замедленной флуоресценции.
В приборе импульсы красного света
миллисекундной длительности и интенсивности, способной запустить процесс
фотосинтеза, подаются от двух мощных светодиодов. Они возбуждают ЗФ, которая
регистрируется в виде кривых затухания в миллисекундные темновые промежутки.
Время световых импульсов и промежутков между ними устанавливаются оператором
перед началом работы. Быстрая флуоресценция, возбуждается от низкоинтенсивного
светодиодного источника синего света, который включается в каждый второй
темновой промежуток между импульсами красного света. Значение БФ вычисляется
путем попарного вычитания из величины интенсивности свечения на момент подачи
импульса синего света, величины свечения (ЗФ), регистрируемой в предыдущий
темновой интервал. Для предотвращения «ослепления» фотоумножителя он
автоматически «запирается» на период подачи каждого импульса красного света с
помощью электронного модулятора. После окончания очередного светового импульса
происходит «отпирание» ФЭУ для регистрации флуоресценции исследуемого
растительного объекта.
Перед началом работы в приборе
размещаются кюветы с образцами, устанавливается режим измерения БФ и ЗФ, и
загружается программный модуль управления прибором. После этого производится
его запуск. Измерение свечения образцов во всех кюветах выполняется в
автоматическом режиме. Включив двигатель вращения кассеты с кюветами, прибор,
ориентируясь на показания датчиков ее положения, находит т.н. «нулевую» кювету,
а затем поочередно выполняет замеры свечения всех других загруженных кювет.
Пустые ячейки кассеты пропускаются прибором. Снятые данные от очередной кюветы
сразу передаются на управляющий компьютер и после обработки высвечиваются на
экране. Для устранения свечения самой кюветы, а также темнового тока, из
значения сигналов кювет вычитается сигнал «нулевой» кюветы. При работе с
суспензиями водорослей или хлоропластов, в нее заливается такой же объем среды,
а в случае анализа «сухих» образцов, она оставляется пустой.
Измерение свечения каждой кюветы
проводится для двух заранее установленных световых и временных режимов, условно
обозначенные как «высокий свет» и «низкий свет». В обоих режимах флуоресценция
образцов регистрируется в виде заданного массива точек для каждой кривой
затухания. В связи с большим объемом получаемой информации, характер затухания
свечения представляется в виде двух усредненных точек - из начала и конца
снятого массива для каждой кривой. При этом на «высоком» свету уровень свечения
отображается начальными точками кривых затухания, а на «низком» - конечными.
Процесс измерения свечений на каждом
свету включает три серии рабочих циклов (рис. 1): серия №1 - измерительный и
действующий света не включены; серия №2 - включен только измерительный свет;
серия №3 - включены оба источника света. Количество рабочих циклов в каждой
серии задается оператором. После установки очередной кюветы сначала следуют
серии замеров на «высоком» свету, затем, через 1 секунду, - на «низком» и
производится перевод кюветы. Возможно проводить измерения свечения каждой
кюветы многократно при работе в режиме нескольких «кругов» вращения кассеты с
кюветами.
Снятые данные и параметры их замера
могут быть сохранены в виде отдельных файлов на управляющем компьютере. При
необходимости они могут быть загружены и просмотрены программой Foton или Microsoft Excel.
В качестве показателей ЗФ взяты
значения амплитуд быстрой (миллисекундной) и медленной (секундной) компонент
затухания послесвечения, а так же их отношение - относительный параметр
замедленной флуоресценции [29].
Результаты статистически обработаны
по стандартным методикам [22].
3. Результаты
исследований
.1 Зависимость ОПЗФ хвои
ели сибирской от района произрастания растений
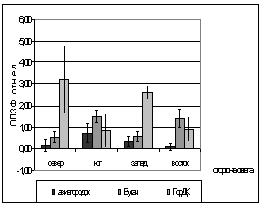
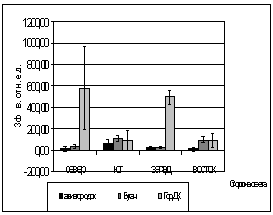
Были проведены опыты, на их
основании можно сделать следующие заключения. Сравнивая площадку №3 (на
рисунках ГорДк) и площадку №2 (на рисунках Авиагородок), мы убеждаемся в их
различии. Средние ОПЗФ на третьей площадке выше, чем средние ОПЗФ с площадки
№2.
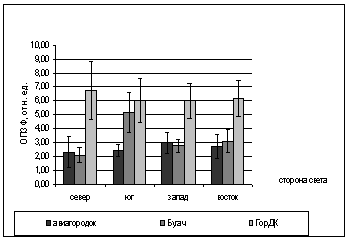
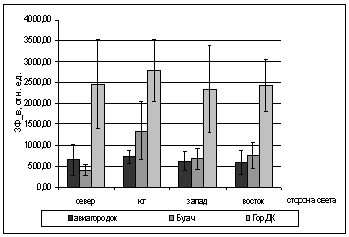
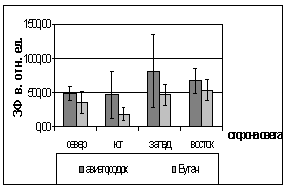
А.
БРис. 2. Зависимость показателей
флуоресценции хвои ели от стороны света и места произрастания деревьев (март
2005).
А. - ОПЗФ; Б. - ЗФ_в.
 А.
А.
 Б.
Б.
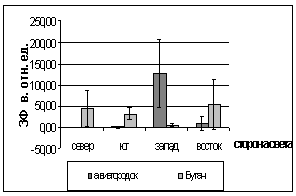
Рис. 3. Зависимость показателей
флуоресценции хвои ели от стороны света и места произрастания деревьев (апрель
2005).
А. - ОПЗФ; Б. - ЗФ_в.
Вероятно, это объясняется различием
атмосферного загрязнения районов. На первой площадке средние ОПЗФ так же ниже
показаний с третьей площадки, это можно объяснить тем, что не далеко находится
КМЗ, но средний поток автотранспорта, что облегчает экологическую обстановку
данной площадки.
В целом, по трем площадкам, опираясь
на собранную информацию, можно сделать вывод, что площадка в районе р. Бугача
находится в более чистом районе, чем площадка в Академгородке, а на ГорДК
деревья испытывают влияние самого большого атмосферного загрязнения.
 А.
А.
 В.
В.
 Б.
Б.
 Г.
Г.
Рис. 4. Зависимость параметров
флуоресценции хвои ели от стороны света и места произрастания деревьев.
А. - ОПЗФ, январь 2005; Б. - ЗФ_в,
январь 2005;
В. - ОПЗФ, февраль 2005; Г. - ЗФ_в,
февраль 2005.
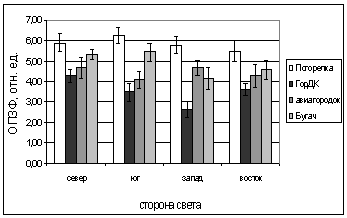
С июня по сентябрь 2005
дополнительно взяли еще одну площадку (д. Погорелка), неподверженную действию
факторов, присутствующих в городской среде. Это позволяет более достоверно
выявить степень угнетения хвойных на остальных площадках. Легко прослеживается
(рис 5 - 6) подтверждение сделанного нами предположения, что в чистом районе,
свободном от выхлопных газов, пыли и других веществ, являющихся следствием деятельности
промышленных центров, растения чувствуют себя гораздо комфортнее. Их ритмы и
циклы не нарушаются, в то время как, в городской среде, хвойные позже уходят в
покой и раньше из него выходят.
 А.
А.
 В.
В.
 Б.
Б.
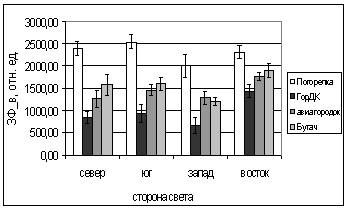 Г.
Г.
Рис. 5. Зависимость параметров
флуоресценции хвои ели от стороны света и места произрастания деревьев.
А. - ОПЗФ, июнь 2005; Б. - ЗФ_в,
июнь 2005;
В. - ОПЗФ, июль 2005; Г. - ЗФ_в,
июль 2005.
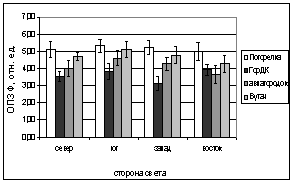 А.
А.
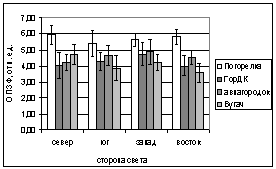 В.
В.
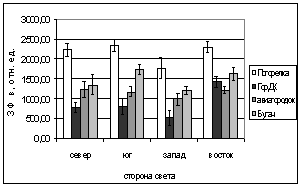 Б.
Б.
 Г.
Г.
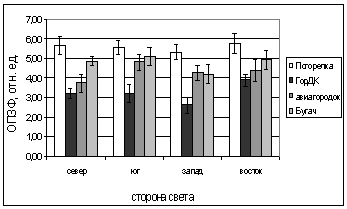
Рис. 6. Зависимость параметров
флуоресценции хвои ели от стороны света и места произрастания деревьев.
А. - ОПЗФ, август 2005; Б. - ЗФ_в,
август 2005;
В. - ОПЗФ, сентябрь 2005; Г. - ЗФ_в,
сентябрь 2005.
3.2 Зависимость ОПЗФ
хвои от стороны света
биоиндикатор загрязнение
сосна воздух
Для изучения высокой вариабельности
результатов опытов бала прослежена зависимость ОПЗФ хвои ели от стороны света.
Прослеживая зависимость ОПЗФ хвои от
стороны света, видно, что на первой площадке показатели преимущественно выше с
южной стороны. На второй площадке наиболее активна хвоя с западной стороны.
Анализируя третью площадку, мы видим, что хвоя с северной стороны света уходит
в менее глубокий зимний покой, чем хвоя с других сторон света.
Такие различия на всех трех
площадках можно объяснить ветром, дующим с разных сторон света, и расположением
промышленных предприятий.
На первой площадке источники,
загрязняющие атмосферу, находились только с западной и южной стороны (КМЗ и
движение автотранспорта), потому что площадка находится на окраине города и с
северной и восточной сторон начинаются поля. Другие две площадки находились
непосредственно в городе. На рисунках c 6 по 9 хорошо видно, что показатели второй площадки намного выше
с западной стороны света, это можно объяснить тем, что с западной стороны
находится крупный перекресток и три автобусные остановки, ветер постоянно
западный. Подобная ситуация на третьей площадке (рис. 2 и 3), объясняется таким
же близким расположением перекрестка и остановок с запада, автобусных остановок
с севера, ветер северо-западный.
На контрольной площадке сложно проследить
какую-либо зависимость ОПЗФ хвои от стороны света, это легко объяснить тем
фактом, что экспериментальную площадку не окружают очевидные источники
загрязняющих веществ.
Дисперсионный анализ говорит о том,
что зависимости ОПЗФ хвои ели от стороны света нет (см. приложение табл. 1 -
3).
3.3 Оценка состояния
зимнего покоя деревьев ели на исследуемых площадках
Для работы с хвоей, закончившей
полный годовой цикл развития, образцы отбирали в зимний период. В это время
года деревья находятся в состоянии зимнего покоя, и по показателям ЗФ и ОПЗФ
можно судить о глубине их покоя, регистрируя показания ЗФ хвои, не выведенной
из состояния покоя.
Было обнаружено, что глубина покоя
существенно различается в зависимости от района. Так, в самом загрязненном
районе (площадка №3) значения ОПЗФ и ЗФ_в значительно выше, чем в других
районах, что говорит о неглубоком зимнем покое. Самые низкие показатели
наблюдаются на второй площадке, испытывающей преимущественно влияние выхлопов
автотранспорта, и находящейся дальше остальных площадок от промышленных
предприятий.
Прослеживая показания от января к
апрелю, видно, что ближе к весне на всех площадках показания ОПЗФ и ЗФ_в
растут. Видимо, деревья с первой и второй площадки, наиболее чистые и
находящиеся в более глубоком покое, чем с третьей площадки, начинают догонять
по активности деревья с последней площадки.
Полученные результаты говорят о том,
что глубина состояния зимнего покоя обратнопропорциональна степени атмосферного
загрязнения.
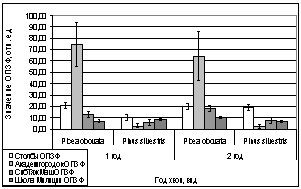
.4 Динамика выхода из
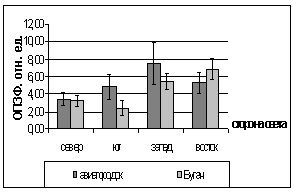
состояния зимнего покоя сосны обыкновенной и ели сибирской
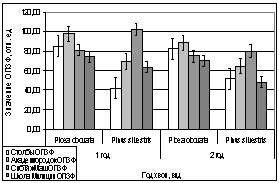
Сравнивая показания с трех пробных
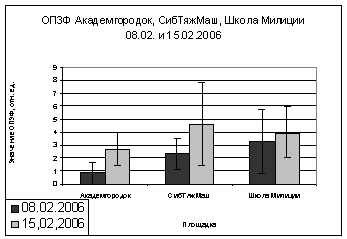
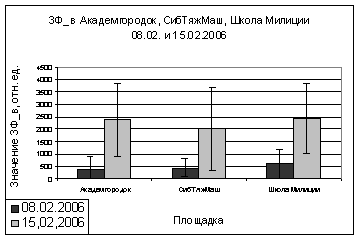
площадок (Академгородок, СибТяжМаш и Школа милиции) в феврале 2006 года, видны
различия. На Школе Милиции показания ОПЗФ и ЗФ_в до экспозиции выше, остальных,
самые маленькие на площадке Академгородок (рис. 7). При статистической
обработке достоверными по площадкам оказались только показания ОПЗФ снятые 8
февраля, остальные не достоверны ни по площадкам, ни по деревьям (см.
приложение табл. 4, 5). Очевидно, в данном случае повлияли индивидуальные
особенности деревьев и площадок. Можно сделать вывод, что в данный период (при
резких колебаниях температуры) использовать сосну обыкновенную в качестве
биоиндикатора не целесообразно.
 А.
А.
 Б.
Б.
Рис. 7. Зависимость
параметров флуоресценции хвои сосны от места расположения площадки (февраль
2006). До и после экспозиции.
А. - ОПЗФ; Б. - ЗФ_в.
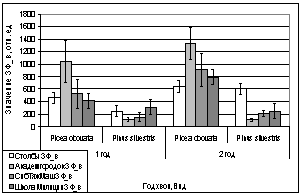
Но анализируя показания
ОПЗФ и ЗФ_в в последующих замеров, видна обратная картина: самые высокие
значения ОПЗФ и ЗФ_в до и после экспозиции на площадке Академгородок, на
СибТяжМаш значительно ниже, а у деревьев со Школы Милиции самые низкие
показания (рис. 8). Достоверность этих показаний ОПЗФ и ЗФ_в подтверждает
статистическая обработка (см. Приложение табл. 6, 7). Это можно объяснить
температурными перепадами в конце февраля.
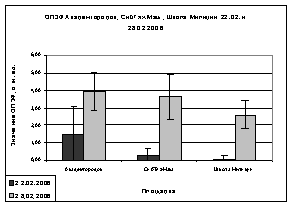 А.
А.
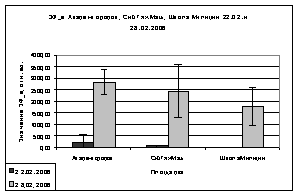 Б.
Б.
Рис. 8. Зависимость
параметров флуоресценции хвои сосны от места расположения площадки (февраль
2006). До и после экспозиции.
А. - ОПЗФ; Б. - ЗФ_в.
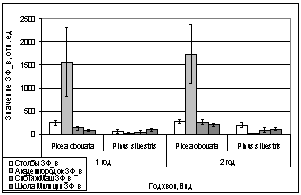
В марте и апреле
показания ОПЗФ и ЗФ_в снова меняют свою направленность - значения
Академгородка, полученные в марте, меньше остальных, на Школе милиции вновь
«вырастают» (рис. 9). В апреле на площадке Академгородок показания ОПЗФ и ЗФ_в
самые высокие, СибТяжМаш меньше и самые низкие на площадке Школа милиции (рис.
10). Это можно так же объяснить сильными перепадами температуры.
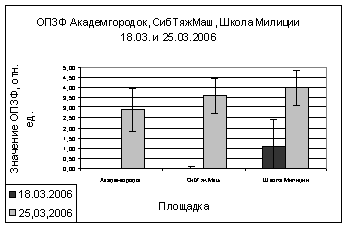 А.
А.
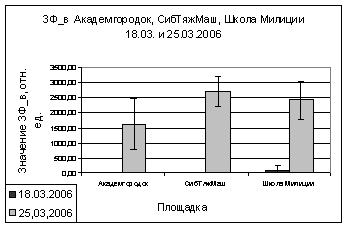 Б.
Б.
Рис. 9. Зависимость
параметров флуоресценции хвои сосны от места расположения площадки (март 2006).
До и после экспозиции.
А. - ОПЗФ; Б. - ЗФ_в.
Если просмотреть все
рисунки (рис. 7 - 10), можно увидеть закономерности динамики выхода деревьев из
состояния покоя. От января к апрелю показания ОПЗФ и ЗФ_в после экспозиции
становятся выше.
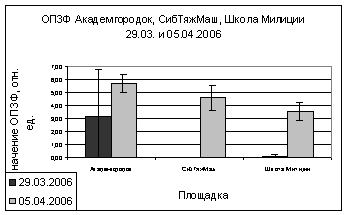 А.
А.
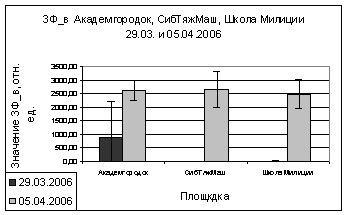 Б.
Б.
Рис. 10. Зависимость
параметров флуоресценции хвои сосны от места расположения площадки (апрель
2006). До и после экспозиции
А. - ОПЗФ; Б. - ЗФ_в.
Как уже упоминалось
выше, показания проведенных опытов были статистически обработаны. Проведен
двухфакторный дисперсионный анализ, в качестве одного фактора взяли
индивидуальные особенности деревьев, которые возможно, могли повлиять на
результаты исследований и ввести в заблуждение, а в качестве второго -
различные условия на площадках, на которые делается основной акцент. Результаты
говорят о том, что индивидуальные особенности деревьев не влияют на результаты
опытов, за исключением первых измерений (о них упоминалось в начале данной
главы). Достоверно, расположение площадок влияет на показатели ОПЗФ и ЗФ_в.
В следующем 2007 году
дополнительно взяли еще одну площадку, которая находится в заповеднике
«Столбы», и еще один вид хвойных ель сибирская (Picea obovata).
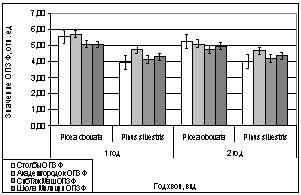 А.
А.
 В.
В.
 Б.
Б.
 Г.
Г.
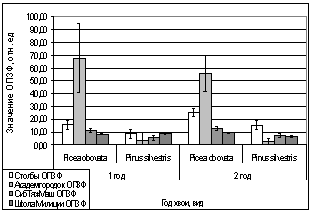
Рис. 11. Зависимость
параметров флуоресценции хвои сосны, ели от места расположения площадки.
А. - ОПЗФ 06.02.2007.
Ночь -10 С, день -1 С.
Б. - ЗФ_в 06.02.2007.
Ночь -10 С, день -1 С.
В. - ОПЗФ 16.02.2007.
Ночь -25 С, день -15 С.
Г. - ЗФ_в 16.02.2007.
Ночь -25 С, день -15 С.
Что касается показаний с
площадки, которая находится в заповеднике «Столбы», то они оказались самыми
большими. Химический анализ показал, что в пробе с данной площадки присутствует
большое количество фторид-иона и / или монокарбоновых кислот, что можно
объяснить достаточно близким расположением большого количества правобережных
заводов и котельных.
Показания ели сибирской
обратнопропорциональны показаниям сосны обыкновенной. Этот факт, возможно,
объясняется меньшей чувствительностью Сосны к кислым газам.
В середине февраля 2007
года температура стала гораздо ниже, чем в начале месяца, что отразилось на
величине показаний флуоресценции хвои с экспериментальных площадок (рис. 12),
они стали меньше, это говорит о том, что процессы метаболизма деревьев снизили
свою скорость. Однако показания хвои ели сибирской с площадки Академгородка
стали гораздо выше. Статистический анализ показал, что эти данные оказались
недостоверными в силу большой вариабельности (см. приложение табл. 12, 13). Это
явление можно объяснить аномально теплой зимой. Ель уходит в состояние покоя
позже сосны и менее глубоко, поэтому реакция на температурные аномалии у
экспериментальных видов разная.
К концу месяца
температурный режим не изменился, ситуация остается прежней, показания ели в
Академгородке такие же высокие (рис. 12). Показания ОПЗФ сосны на
экспериментальных площадках несколько ниже, показаний ели, что объясняет
меньшая готовность к состоянию покоя Pinus sylvestris.
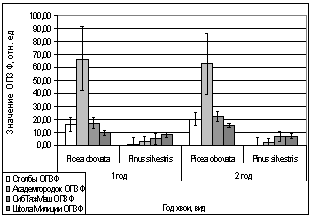 А.
А.
 В.
В.
 Б.
Б.
 Г.
Г.
Рис. 12. Зависимость
параметров флуоресценции хвои сосны, ели от места расположения площадки.
А. - ОПЗФ 22.02.2007.
Ночь -25 С, день -18 С;
Б. - ЗФ_в Милиции
22.02.2007. Ночь -25 С, день -18 С;
В. - ОПЗФ 01.03.2007.
Ночь -25 С, день -15 С;
Г. - ЗФ_в 01.03.2007.
Ночь -25 С, день -15 С.
В первой половине марта,
не смотря на повышение температуры, значения ОПЗФ так же не превышают 10 отн.
ед. Видимо, сезонные биоритмы, продолжительность светового дня не позволяют
деревьям, у которых брали образцы, ускорить свои метаболические процессы,
«распаковать» больше тилакоидов хлоропластов. Опять можно проранжировать
экспериментальные площадки (рис. 13), как в предыдущем месяце от меньшего загрязнения
атмосферы района к большему - Академгородок, СибТяжМаш, Школа милиции со
«Столбами». Исключение составляет опять же ель на площадке Академгородка.
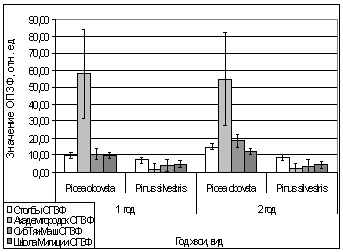 А.
А.
 В.
В.
 Б.
Б.
 Г.
Г.
Рис. 13. Зависимость
параметров флуоресценции хвои сосны, ели от места расположения площадки.
А. - ОПЗФ 10.03.2007.
Ночь -12 С, день -5 С;
Б. - ЗФ_в 10.03.2007.
Ночь -12 С, день -5 С;
В. - ОПЗФ 16.03.2007.
Ночь -4 С, день 0 С;
Г. - ЗФ_в 16.03.2007.
Ночь -4 С, день 0 С.
Уже при температурах
близких к нулю, установившихся ближе к концу марта, ель начинает потихоньку
выходить из состояния глубокого покоя, о чем говорят несколько поднявшиеся
параметры флуоресценции (рис. 14).
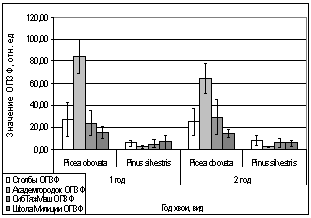 А.
А.
 В.
В.
 Б.
Б.
 Г.
Г.
Рис. 14. Зависимость параметров
флуоресценции хвои сосны, ели от места расположения площадки.
А. - ОПЗФ 22.03.2007.
Ночь -4 С, день +2 С;
Б. - ЗФ_в 22.03.2007.
Ночь -4 С, день +2 С;
В. - ОПЗФ 31.03.2007.
Ночь -8 С, день -2 С;
Г. - ЗФ_в 31.03.2007.
Ночь -8 С, день -2 С.
В конце марта - начале
апреля, при тех же температурных условиях, близких к нулю, начинает выходить из
покоя сосна. Двухнедельную задержку сосны от ели также можно объяснить ее
меньшей морозоустойчивостью и холодостойкостью. Показания флуоресценции
образцов хвои сосны примерно одинаковые (рис. 14). А по показаниям ОПЗФ и ЗФ
хвои ели по-прежнему и в весенний период можно судить об экологической
обстановке, складывающейся в районах экспериментальных площадок.
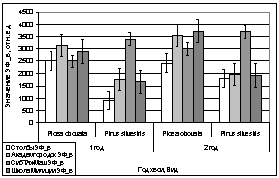
В первой половине
апреля, показания флуоресценции образцов хвои обоих видов начинают ровняться,
это видно не рисунке с 15. А, Б. Отношения показаний ЗФ и ОПЗФ остаются
прежними, как относительно видов хвойных, так и относительно площадок.
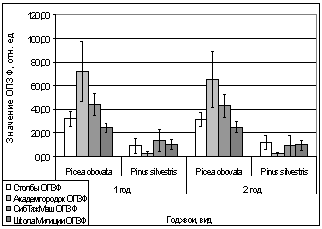
 А.
А.
 В.
В.
 Б.
Б.
 Г.
Г.
Рис. 15. Зависимость
параметров флуоресценции хвои сосны, ели от места расположения площадки.
А. - ОПЗФ 10.04.2007.
Ночь +2 С, день +12 С;
Б. - ЗФ_в 10.04.2007.
Ночь +2 С, день +12 С;
В. - ОПЗФ 19.04.2007.
Ночь +6 С, день +10 С;
Г. - ЗФ_в 19.04.2007.
Ночь +6 С, день +10 С.
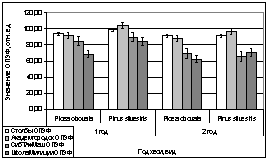
Во второй половине
апреля 2007 года деревья окончательно выходят из состояния зимнего покоя,
устанавливается более стабильная плюсовая температура. На 15 рисунке наиболее
хорошо видно, что по показаниям флуоресценции хвои ели в это время года
проводить биоиндикационные исследования нельзя в силу их нестабильности.
3.5 Влияние
предварительного охлаждения побегов сосны собранных в районах разной степени
загрязнения
Летом 2006 года были
проведены три опыта, направленные на изучение влияния предварительного
охлаждения до отрицательных температур побегов сосны собранных в летнее время в
районах разной степени загрязнения. Первый опыт проводился в период с 14 по 24
июня 2006 года. Как видно на рисунке (рис. 16), при экспозиции побегов в
течение десяти суток при температуре -2ºС
показания ОПЗФ и ЗФ_в снизились незначительно. Оперируя этой информацией, можно
сделать вывод, что при отрицательных близких к нулю температурах уровень
метаболизма снижается, растения уходят в неглубокое состояние покоя. Нужно
отметить, что образцы, собранные на площадке Школа Милиции, менее готовы
перейти в состояние покоя, чем из Академгородка, это можно объяснить разной
экологической обстановкой в наблюдаемых районах. Видна обратная
пропорциональность: в более чистом районе, Академгородке, показания ЗФ_в
образцов в течение опыта становились меньше, а значения ЗФ_в с площадки Школа
Милиции увеличились. У образцов из более чистого района, где у деревьев
показатели ЗФ более высокие и, соответственно, накоплено больше веществ,
необходимых для перехода в состояние покоя, показания ЗФ_в хвои после
экспозиции снижаются. А хвоя со Школы милиции наоборот после десяти суток
экспозиции при -2°С, становится активнее. Такую реакцию можно объяснить
стрессовой теорией, в которой говорится о том, что при стрессовой ситуации
клетка мобилизируется, метаболизм активируется до предела (в этой ситуации
клетка либо переживает стресс, либо погибает).
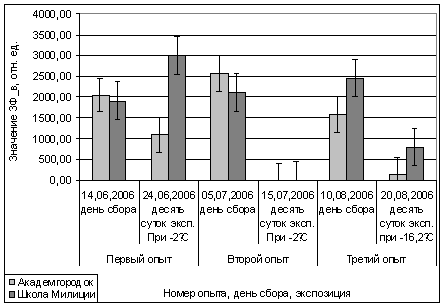
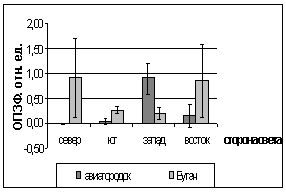
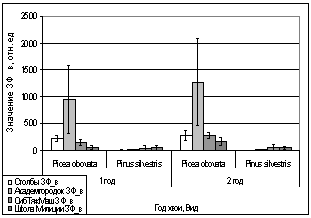
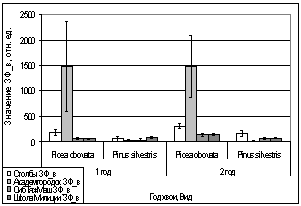
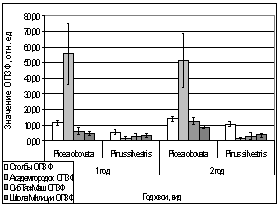
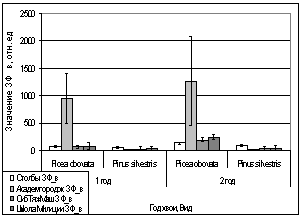
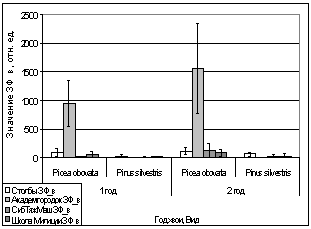
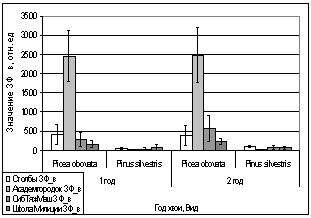
Рис. 16. Значение ЗФ_в
хвои второго года жизни взятой в разных районах
Второй опыт проводился в
период с 5 по 15 июля 2006 года. Использовались те же условия, что и в первом
опыте, отрицательная температура близкая к нулю. Анализируя рисунок 16, видим,
что результаты существенно изменились по сравнению с предыдущим опытом.
Практически все показания флуоресценции хлорофилла образцов равны нулю, не
зависимо от района, в котором находилась площадка. Это говорит о том, что в
данном опыте растения ушли в более глубокое состояние покоя, чем в предыдущем
опыте, так как деревья к этому времени накопили достаточно веществ необходимых
для погружения в покой. Нужно отметить, что после медленного нагревания до
комнатной температуры образцы из 1 и 2 опыта восстанавливали свой метаболизм.
Результаты третьего
опыта видны на рисунке 16. Опыт проводился с 10 по 20 августа 2006 года.
Собранную хвою поместили на мороз (-16,2°С). Образцы после экспозиции ушли в
глубокий покой и их фотосинтетический аппарат после размораживания не
восстановился. Это говорит о необходимости прохождения фазы осенней
фотопериодической реакции.
Выводы
. Динамика
относительного показателя замедленной флуоресценции хлорофилла ели сибирской и
сосны обыкновенной различна в районах с разным уровнем загрязнения воздуха. Чем
меньше загрязнение атмосферы, тем ниже показатели ОПЗФ хвои в зимний период и,
соответственно, более глубокое состояние зимнего покоя. В летний период,
наоборот, более высокие значения ОПЗФ наблюдаются в менее загрязненных районах.
. Достоверных отличий (р
= 0,95) между параметрами ЗФ хвои сосны и ели, взятой из частей кроны с разных
сторон света, не прослеживается.
. В период вынужденного
покоя биоиндикационные возможности хвойных во многом зависят от климатических
особенностей года. В период аномально теплых зим деревья с различной скоростью
выходят из состояния зимнего покоя, что обусловлено биолог8ическими
особенностями растений и затрудняет проведение биоиндикационных исследований.
Ввиду высокой вариабельности в этот период использование сосны и ели в
биоиндикационных целях нежелательно. Оптимально использовать хвойные в качестве
биоиндикаторов с конца марта до октября.
. При охлаждении побегов
сосны обыкновенной, собранных в летние месяцы, до отрицательных близких к нулю
температур (-2°С) в течение 10 суток было отмечено, что в начале лета изменения
параметров ЗФ незначительны, но хвоя из условно чистого района имеет большую
готовность к переходу в состояние покоя. Со второй половины лета при охлаждении
в течение 10 суток образцы и из чистого и из загрязненного района полностью
переходят в состояние временного покоя. При воздействии более низкими
температурами (-16°С) и экспозиции в течение 10 суток, была отмечена гибель хвои с
обоих площадок, что объясняется отсутствием необходимых изменений, которые в
природных условиях протекают в ходе осенней фотопериодической реакции.
Литература
1. Биоиндикация загрязнений наземных экосистем / Под ред. Р.
Шуберта. - М: Мир, 1988. С. 9-29.
2. Веселовский В.А., Веселова Т.В. Люминесценция растений:
Теоретические и практические аспекты. - М.: Наука, 2010. уп - 200 с.
. Гаевский Г.А., Сорокина Г.А., Гольд В.М. Сезонные
изменения фотосинтетического аппарата древесных растений // Физиология
растений. - 2010. - Т. 40.
. Гаевский Н.А. Моргун В.Н. Использование переменной и
замедленной флуоресценции хлорофилла для изучения фотосинтеза растений /
Физиология растений. - 2007.- Т. 40. - С. 136-145.
. Григорьев Ю.С., Бучельников М.А. Биоиндикация загрязнений
воздушной среды на основе замедленной флуоресценции хлорофилла листьев и
феллодермы деревьев // Экология, 2009. №4. С. 303-305, 273-275.
. Григорьев Ю.С., Пахарькова Н.В. Влияние техногенного
загрязнения воздушной среды на состояние зимнего покоя сосны обыкновенной //
Экология. - 2006. - №6. - С. 471 - 473.
. Гудериан Р. Загрязнение воздушной среды. - М.: Мир, 1979.
. Загрязнение воздуха и жизнь растений // под ред. М.
Трешоу. - Л. Гидрометиздат, 1988. - с. 535.
. Зверева Г.Н., Трунова Т.И. Зависимость морозостойкости
озимой пшеницы от синтеза белка во время закаливания // Физиол. растений. -
1985. - Т.32, №5. - С. 976 - 982.
. Изменения жирнокислотного состава липидов сосны
обыкновенной в течение годичного цикла / И.Л. Фуксман, М.К. Ильинова, Л.А.
Сауккнен, А.А. Степанов // Химия древесины. - 1986. - №3. - С. 101 - 108.
. Илькун Г.М. Газоустойчивость растений // Вопросы экологии
и физиологии. Киев, Наукова думка, 1971. - 145 с.
. Катц М. некоторые аспекты физической и химической природы
атмосферных загрязнений. - В кн.: загрязнение атмосферного воздуха. - Всемирная
организация здравоохранения. Женева, 1962.
. Климов С.В. Биоэнергетическая концепция устойчивости
растений к низким температурам // Успехи совр. биол. - 1997. - Т.117, №2. - С.
133 - 154.
. Комбияйнен В.К., Болондинский В.К. водный режим и
фотосинтез сосны в условиях промышленного загрязнения // физиология растений -
2006. Т 42 №3 - с. 86 - 93.
. Конев С.В. Лабильность биологических мембран и
регуляторные процессы. - Минск: Наука и техника, 1987. - 240 с.
. Красавцев О.А. Калориметрия растений при температуре ниже
нуля. - М.: Наука, 1972. - 117.
. Красавцев О.А., Хвалин Н.Н. Незамерзающая вода в
закаленных зачаточных побегов озимых злаков // Физиол. растений. - 1982. -
Т.29. - №3. С. 437 - 446.
. Красинский Н.П. Теоретические основы построения
ассортиментов газоустойчивых растений. - В кн.: Дымоустойчивость растений и
дымоустойчивые ассортименты. Москва - Горький, 1950 - 160 с.
. Кривошеева А.А., Сорокина Г.А., Гольд В.М. влияние
промышленного загрязнения на сезонные изменения содержания хлорофилла в хвое
сосны обыкновенной // Физиология растений. - 2008. - Т. 35.
. Кузина Г.В., Калинина Г.А. Содержание АБК в процессе
прохождения осенней фотопериодической реакции, индукция глубокого покоя и
морозостойкости абрикоса // Физиология растений. - 2006. - Т.40 №3, с. 397 -
405.
. Лакин Г.Ф. Биометрия. - М: Выс. школа. - 1980.
. Мануильская С.В. Роль липидных компонентов мембран
тилакоидов в функционировании фотосинтетического аппарата и адаптации к
условиям внешней среды // Физиология и биохимия культурных растений. - 1987. -
Т. 19. №1, с. 29 - 34.
. Маторин Д.Н., Венедиктов П.С., Рубин А.В. Замедленная
флуоресценция и ее использование для оценки состояния растительного организма /
Изв. АН СССР серия биологическая №4. - 1985. - С. 9-13.
. Мониторинг фонового загрязнения / Под ред. Ю.А. Израэля. -
М.: Знание, 1987. - с. 43-46.
. Мэнинг У.Д., Федер У. Биомониторинг загрязнений атмосферы
с помощью растений. - Л.: Гидрометиоиздат. 1985. - 143 с.
. Николаевский В.С. Биологические основы газоустойчивости
растений. - Новосибирск: Наука, 1979. - 280 с.
. Новицкая Ю.Е., Чикина П.Ф. Азотный обмен сосны на севере.
- Л.: Наука, 1980. - 166 с.
. Пахарькова Н.В. Замедленная флуоресценция хлорофилла
хвойных в условиях техногенного загрязнения атмосферы / Автореферат диссертации
на соискание ученой степени кандидата биологических наук. - Красноярск, 2009.
. Петров В.В. Лес и его жизнь. - М: Просвещение, 1986. - С.
14-27.
. Пых Ю.А., Малкина-Пых И.Г. Об оценке состояния окружающей
среды. Подходы к проблеме // Экология, 2006. №5. С. 323-329.
. Регуляторы роста растений. Под ред. В. Кефели. - М.:
Знание, 1988. - с. 18 - 30.
. Рожков А.С., Михайлова Т.А. Действие фотосодержащих
эмиссий на хвойные деревья. Новосибирск: Наука, 1989. - С. 156.
. Рост и морозостойкость растений / Т.И. Трунова, Г.В.
Кузина, М.А. Бочарова, Н.В. Астахова // В кн. Рост и морозостойкость растений.
- Новосибирск: Наука, 1988. - С. 133 - 143.
. Рост растений и дифференцировка // под ред. Ф. Уоринг. -
М.: Мир, 1984. - с. 67 - 84.
. Рубин А.Б. Биофизические методы в экологическом
мониторинге // Соросовский образовательный журнал, 2006. Том 6. №4. С. 7-13.
. Рубчевская Л.П. Глицериды камбиальной зоны лиственницы
сибирской: Автореф. дис…канд. хим. наук. - Рига, 1983. - 24 с.
. Самыгин Г.А. На каком из этапов замораживания-оттаивания
внеклеточный лед повреждает клетки // Физиол. раст. - 1988. - Т. 35. - №2. - С.
341 - 348.
. Самыгин Г.А. Причина вымерзания растений. - М.: Наука,
1974 - 192 с.
. Самыгин Г.А. Причины повреждения клеток растений
внеклеточным льдом // Физиол. раст. - 2008. - Т. 41, №4. - С. 614 - 625.
. Сергеев Л.И., Сергеева К.А. Структурно-метаболические
механизмы адаптации древесных растений к неблагоприятным факторам внешней среды
// Сезонные структурно-метаболические ритмы и адаптация древесных растений. -
Уфа: изд. БаФ АН СССР, 1977. - С. 11 - 36.
. Сергеева К.А. Физиологические и биохимические основы
зимостойкости древесных растений. - М.: Наука, 1971. - 174 с.
. Судачкова Н.Е. Метаболизм хвойных и формирование
древесины. - Новосибирск: Наука, 1977. - 230 с.
. Судачкова Н.Е., Семенова Г.П. Белки клеточных стенок
ксилемы древесных растений и их функции в регуляции роста клеток //
Лесоведение. - 1988. - №1. - С. 26 - 32.
. Судачкова Н.Е., Семенова Г.П. К характеристике белков
вегетативных органов кедра сибирского // Материалы конференции по физиологии и
биохимии растений. - Красноярск, 1972. - С. 53 - 57.
. Судачкова Н.Е., Семенова Г.П. Растворимые белки
прикамбиальной зоны Pinus sylvestris L. // Лесоведение. - 1971. - №6. - С. 39 - 44.
. Тарабрин В.П. фитотоксичность органических и неорганических
загрязнений. - Киев, 1986.
. Трунова Т.И. Физиологические и биохимические адаптации
растений к морозу // С.-Х. биол. - 1984. - №6. С. 3 - 6.
. Туманов И.И. Физиология закаливания и морозостойкости
растений. - М.: Наука, 1979 - 352 с.
. Тюрин М.М. Развитие представления о состоянии покоя
древесных растений // Физиология растений. - 1986. - Т. 33, вып. 1. - с. 899 -
905.
. Физиология растений // под ред. В. Артамонова - М.:
Агропромиздат, 2008. - с. 326 - 334.
. Флуориметр для биологического контроля загрязнения
окружающей среды (Фотон-9). Паспорт. Техническое описание. Инструкция по
эксплуатации. Красноярск, 2008.
. Фотосинтез: физиолого-экологические и биохимические
аспекты // под ред. Л. Мокроносова. - М.: Изд-во МГУ, 1992. - с. 42 - 99.
. Ходасевич А.В. Фотосинтетический аппарат хвойных //
Минск: Наука и техника, 1982.-С. 199.
. Чикина П.Ф. Динамика различных форм азота в органах и
тканях сосны // Физиолого-биохимические основы роста и адаптации сосны на
севере. - Л.: Наука, 1985. - С. 57 - 63.
. Шванин С.А., Кирпичникова В.В. Состояние
фотосинтетического аппарата сосны обыкновенной и ели в зонах промышленного
загрязнения при разных микроклиматических условиях. // Физиология растений. -
2008, №1. - с. 107.
. Шихов Н.С. Биохимическая оценка состояния городской среды
// Экология, - №2, - с. 146 - 149.
. Юсуфоф А.Г. Устойчивость растений к неблагоприятным
условиям. - М.: Знание, 1986. - с. 46 - 51.
59. Huttunen S., Korenlampi I., Kolari K. Changes of osmotic
values and come related physiological variables in polluted coniferous needles
// ann/ bot. Fenn. 1981, 18. - p. 63 - 71.
. Levitt J. Responses of plants to environmental stzesses.
New York: Academic Press, 1972 - 697 pp.
. Soikkelis S., Tujvinen T. Damage in mesophyll
ultrastrukture of needles of Norway Spruce in industrial environments in
Central Finland. Ann. Bot. fenn., 1979. - Vol. 16. - N1. - p. 50 - 64.